

Vitamin B12 in the spotlight again
- PMID: 28167430
- PMCID: PMC5540639
- DOI: 10.1016/j.cbpa.2017.01.013
Vitamin B12 in the spotlight again
Abstract
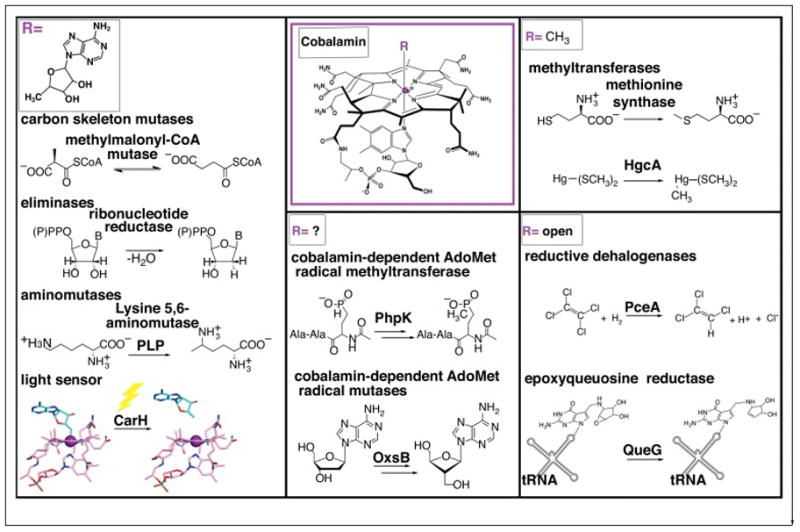
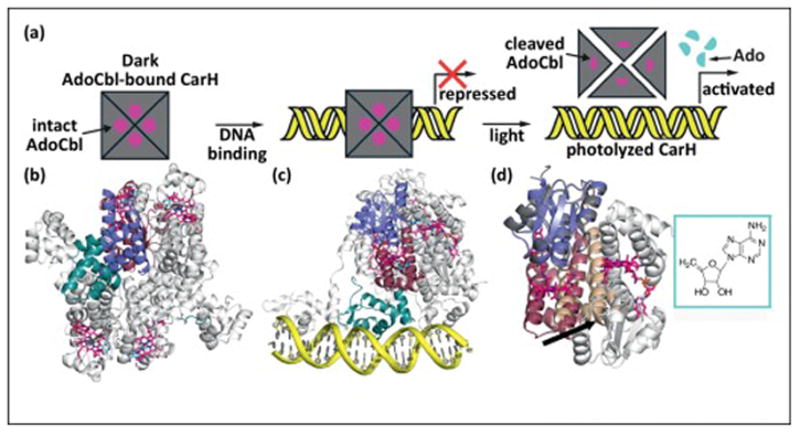
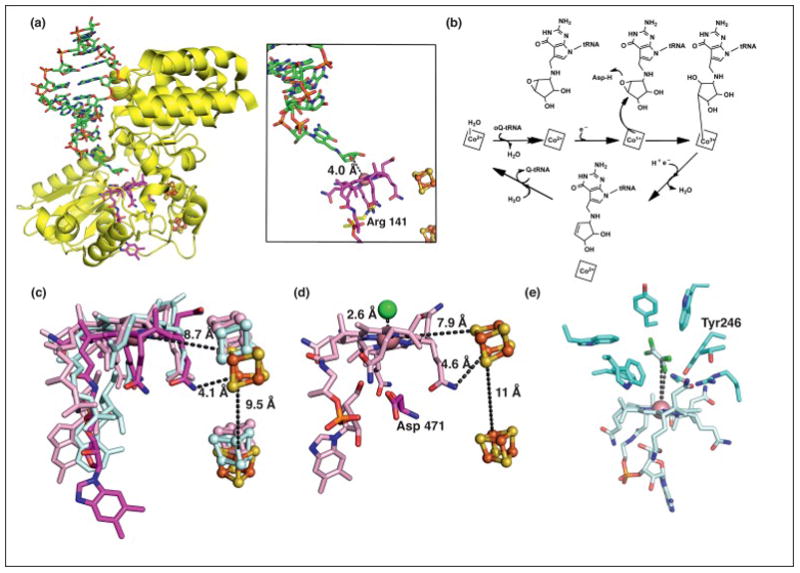
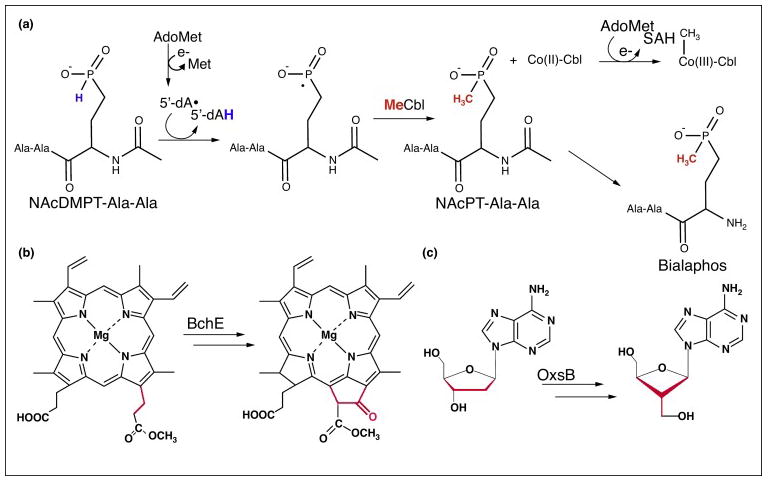
The ability of cobalamin to coordinate different upper axial ligands gives rise to a diversity of reactivity. Traditionally, adenosylcobalamin is associated with radical-based rearrangements, and methylcobalamin with methyl cation transfers. Recently, however, a new role for adenosylcobalamin has been discovered as a light sensor, and a methylcobalamin-dependent enzyme has been identified that is suggested to transfer a methyl anion. Additionally, recent studies have provided a wealth of new information about a third class of cobalamin-dependent enzymes that do not appear to use an upper ligand. They function in reductive dehalogenations and epoxide reduction reactions. Finally, mechanistic details are beginning to emerge about the cobalamin-dependent S-adenosylmethionine radical enzyme superfamily for which the role of cobalamin has been largely enigmatic.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
References
-
- Scott AI, Roessner CA. Biosynthesis of cobalamin (vitamin B(12)) Biochem Soc Trans. 2002;30:613–620. - PubMed
-
- Croft MT, Lawrence AD, Raux-Deery E, Warren MJ, Smith AG. Algae acquire vitamin B12 through a symbiotic relationship with bacteria. Nature. 2005;438:90–93. - PubMed
-
- Padovani D, Labunska T, Banerjee R. Energetics of interaction between the G-protein chaperone, MeaB, and B12-dependent methylmalonyl-CoA mutase. J Biol Chem. 2006;281:17838–17844. - PubMed
-
- Jost M, Cracan V, Hubbard PA, Banerjee R, Drennan CL. Visualization of a radical B12 enzyme with its G-protein chaperone. Proc Natl Acad Sci U S A. 2015;112:2419–2424. This work presents structural characterization of a fusion between an AdoCbl-dependent isobutyryl-CoA mutase and its G-protein chaperone. The structures presented here provide the first insight into the necessary conformational changes for loading a precious molecule of AdoCbl in an enzyme active site. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
