

Comprehensive characterization of neutrophil genome topology
- PMID: 28167501
- PMCID: PMC5322729
- DOI: 10.1101/gad.293910.116
Comprehensive characterization of neutrophil genome topology
Abstract
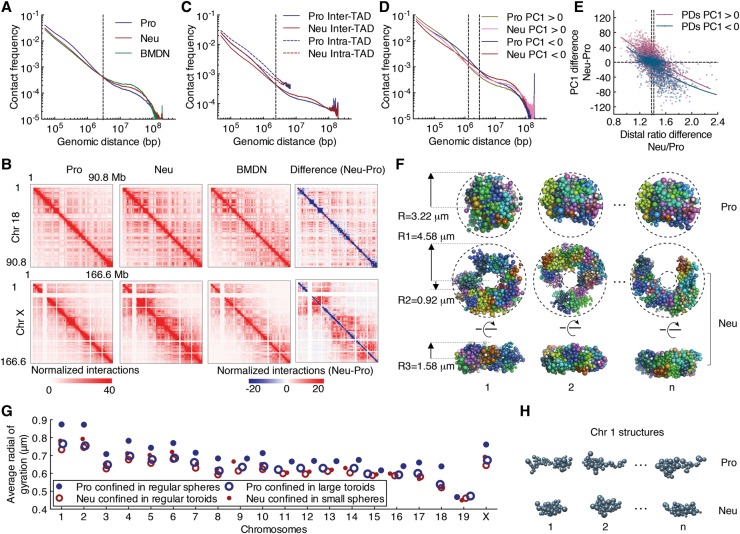
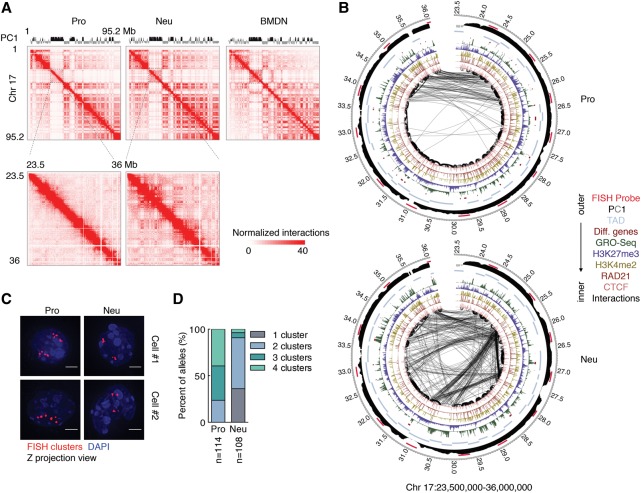
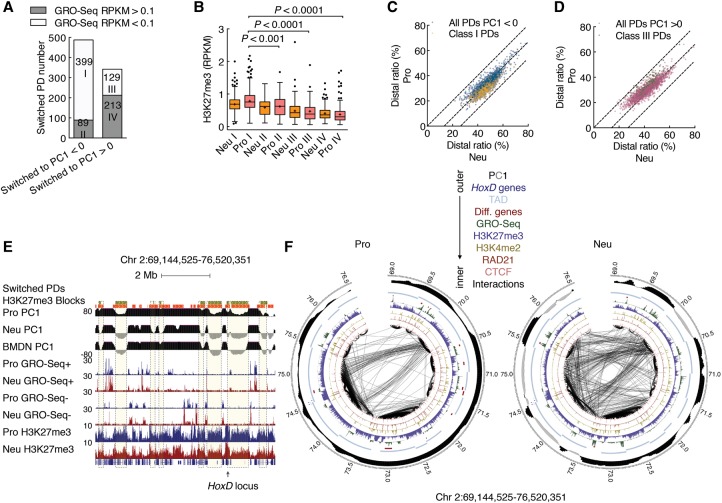
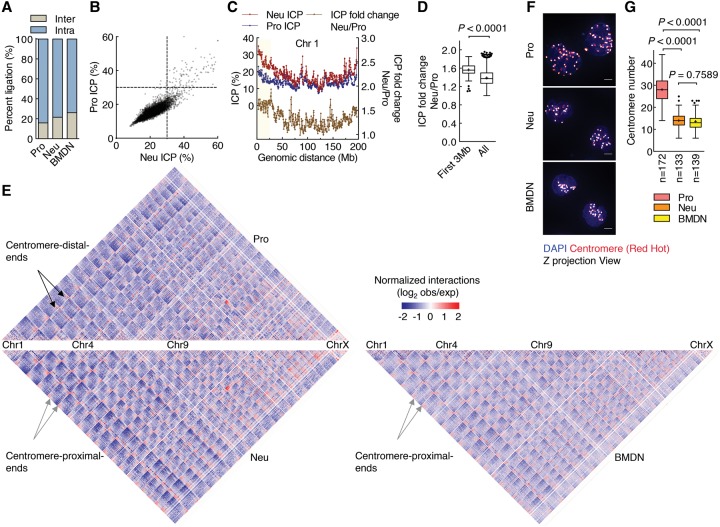
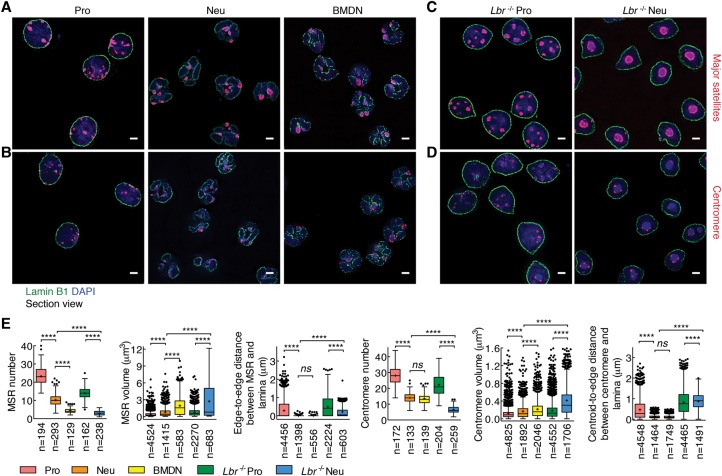
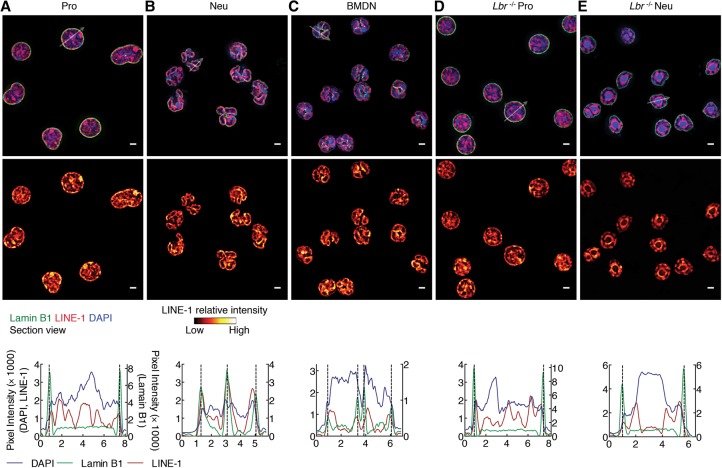
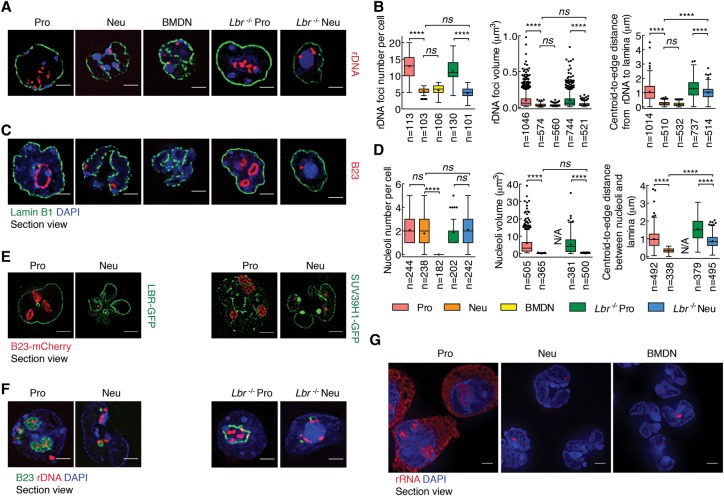
Neutrophils are responsible for the first line of defense against invading pathogens. Their nuclei are uniquely structured as multiple lobes that establish a highly constrained nuclear environment. Here we found that neutrophil differentiation was not associated with large-scale changes in the number and sizes of topologically associating domains (TADs). However, neutrophil genomes were enriched for long-range genomic interactions that spanned multiple TADs. Population-based simulation of spherical and toroid genomes revealed declining radii of gyration for neutrophil chromosomes. We found that neutrophil genomes were highly enriched for heterochromatic genomic interactions across vast genomic distances, a process named supercontraction. Supercontraction involved genomic regions located in the heterochromatic compartment in both progenitors and neutrophils or genomic regions that switched from the euchromatic to the heterochromatic compartment during neutrophil differentiation. Supercontraction was accompanied by the repositioning of centromeres, pericentromeres, and long interspersed nuclear elements (LINEs) to the neutrophil nuclear lamina. We found that Lamin B receptor expression was required to attach centromeric and pericentromeric repeats but not LINE-1 elements to the lamina. Differentiating neutrophils also repositioned ribosomal DNA and mininucleoli to the lamina-a process that was closely associated with sharply reduced ribosomal RNA expression. We propose that large-scale chromatin reorganization involving supercontraction and recruitment of heterochromatin and nucleoli to the nuclear lamina facilitates the folding of the neutrophil genome into a confined geometry imposed by a multilobed nuclear architecture.
Keywords: Lamin B receptor; genome topology; neutrophils; nucleoli; rDNA.
© 2017 Zhu et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
Comment in
-
In search of the mechanism that shapes the neutrophil's nucleus.Genes Dev. 2017 Jan 15;31(2):85-87. doi: 10.1101/gad.296228.117. Genes Dev. 2017. PMID: 28202537 Free PMC article.
References
-
- Apostolou E, Thanos D. 2008. Virus infection induces NF-κB-dependent interchromosomal associations mediating monoallelic IFN-γ gene expression. Cell 134: 85–96. - PubMed
-
- Bickmore WA, Van Steensel B. 2013. Genome architecture: domain organization of interphase chromosomes. Cell 152: 1270–1284. - PubMed
-
- Chow JC, Ciaudo C, Fazzari MJ, Mise N, Servant N, Glass JL, Attreed M, Avner P, Wutz A, Barillot E, et al. 2010. LINE-1 activity in facultative heterochromatin formation during X chromosome inactivation. Cell 141: 956–969. - PubMed
-
- Corbel C, Diabangouaya P, Gendrel A-V, Chow JC, Heard E. 2013. Unusual chromatin status and organization of the inactive X chromosome in murine trophoblast giant cells. Development 140: 861–872. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases