

Identification of a Membrane Protein Required for Lipomannan Maturation and Lipoarabinomannan Synthesis in Corynebacterineae
- PMID: 28167532
- PMCID: PMC5377810
- DOI: 10.1074/jbc.M116.772202
Identification of a Membrane Protein Required for Lipomannan Maturation and Lipoarabinomannan Synthesis in Corynebacterineae
Abstract
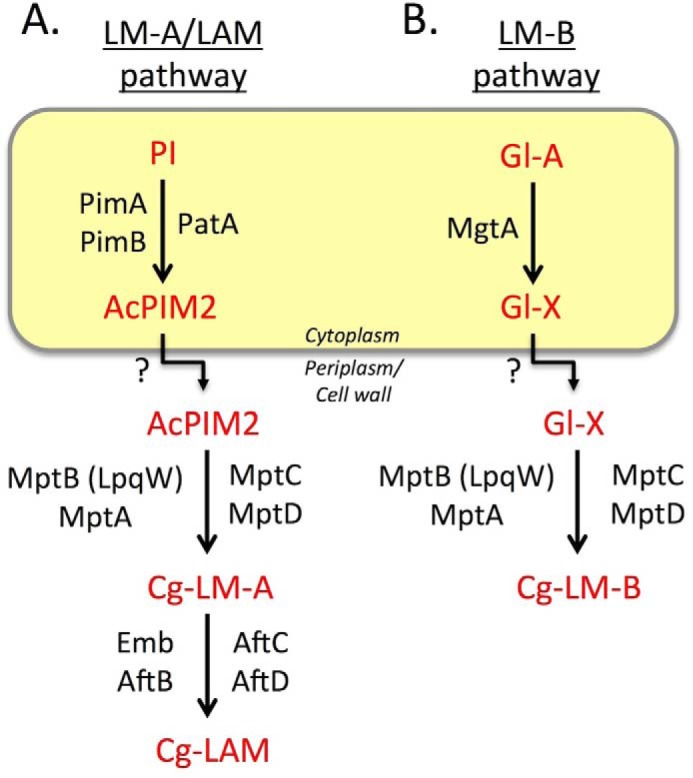
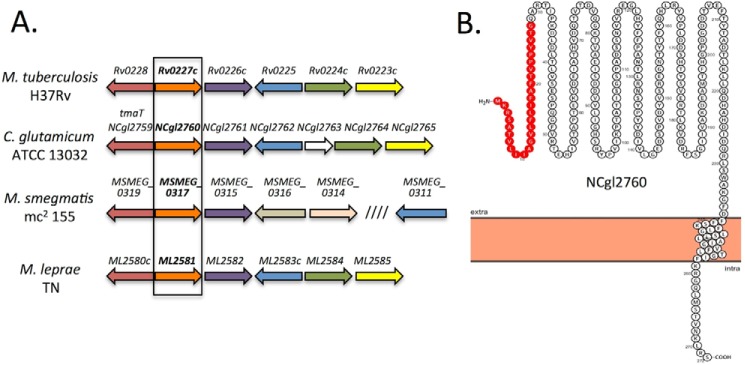
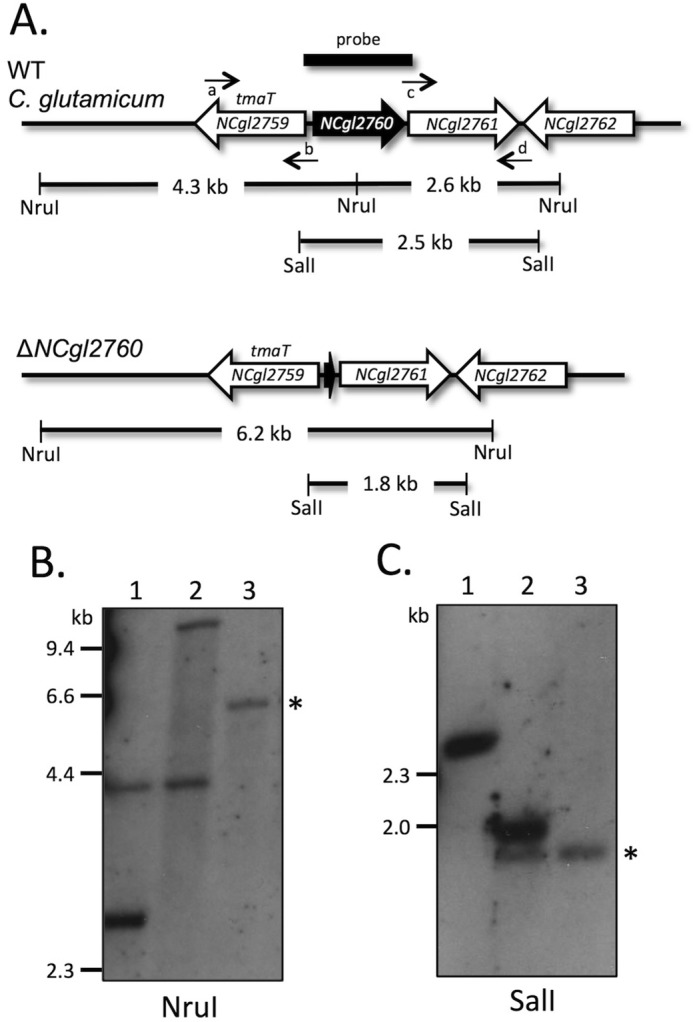
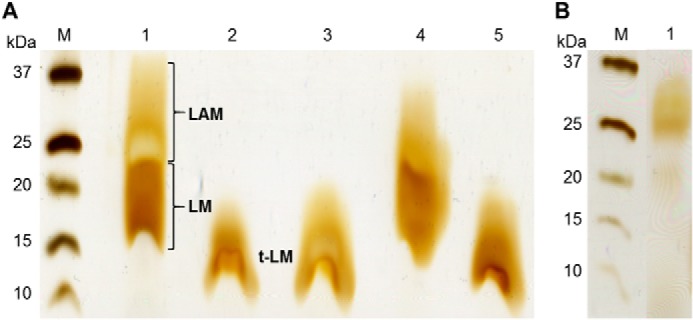
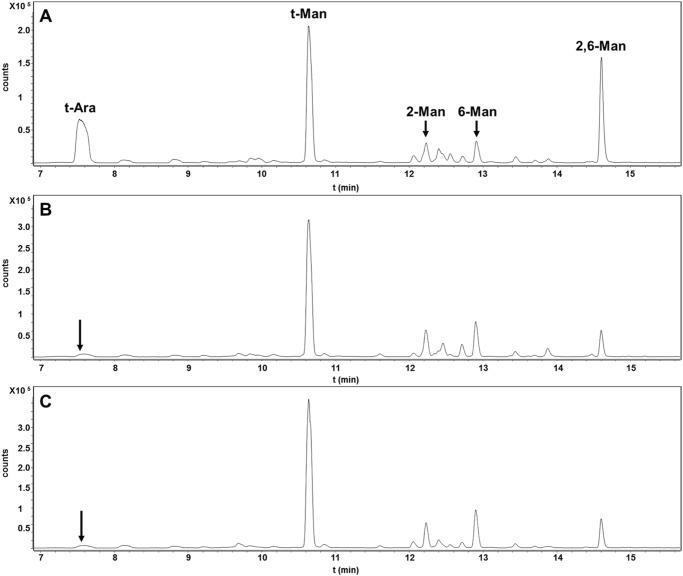
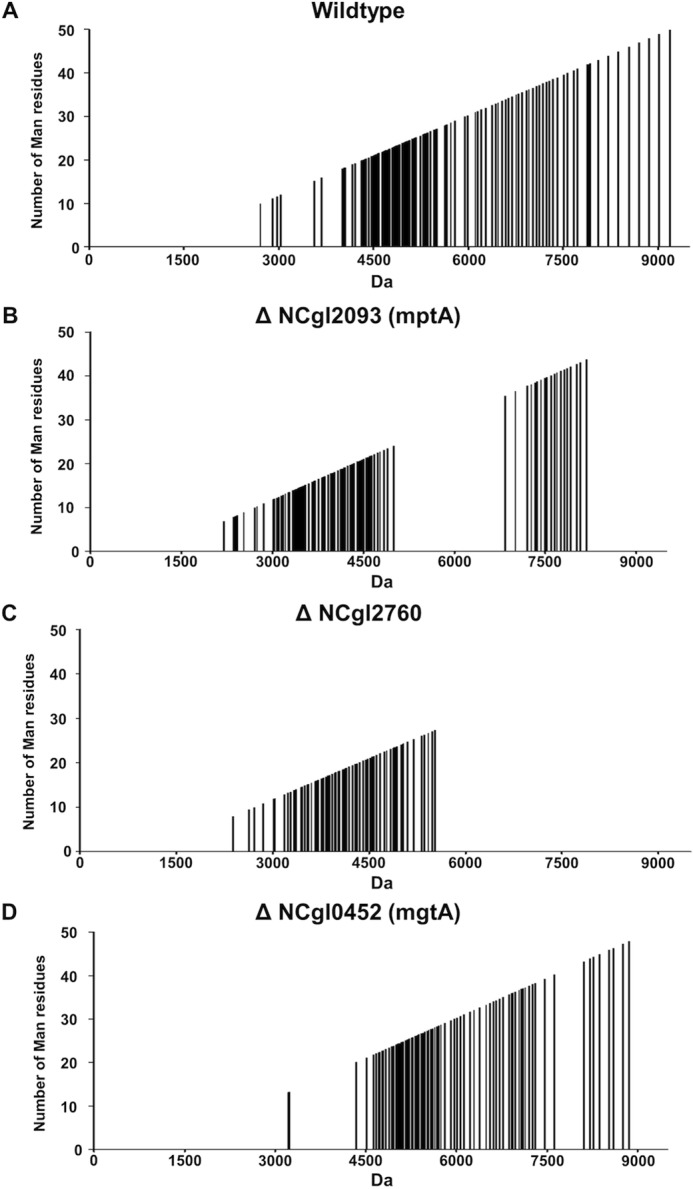
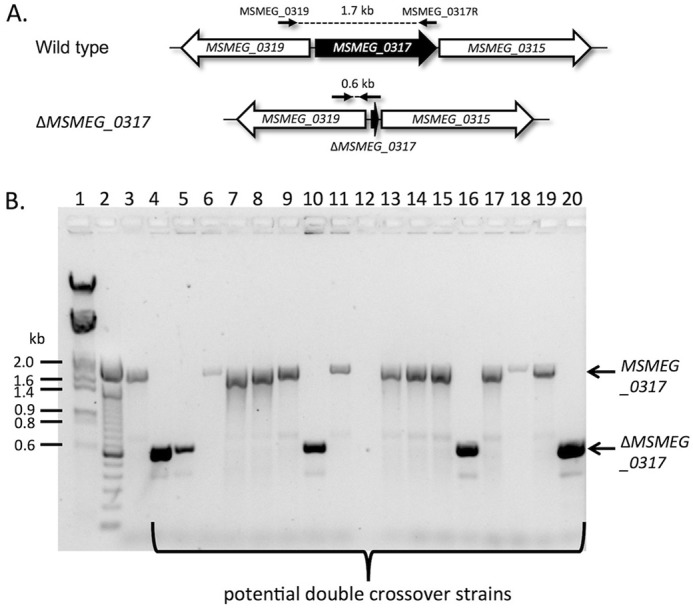
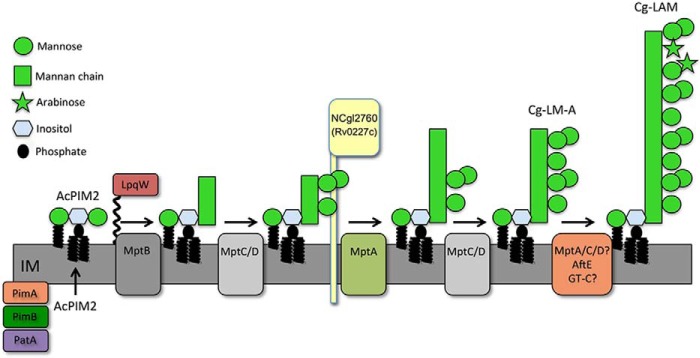
Mycobacterium tuberculosis and related Corynebacterineae synthesize a family of lipomannans (LM) and lipoarabinomannans (LAM) that are abundant components of the multilaminate cell wall and essential virulence factors in pathogenic species. Here we describe a new membrane protein, highly conserved in all Corynebacterineae, that is required for synthesis of full-length LM and LAM. Deletion of the Corynebacterium glutamicum NCgl2760 gene resulted in a complete loss of mature LM/LAM and the appearance of a truncated LM (t-LM). Complementation of the mutant with the NCgl2760 gene fully restored LM/LAM synthesis. Structural studies, including monosaccharide analysis, methylation linkage analysis, and mass spectrometry of native LM species, indicated that the ΔNCgl2760 t-LM comprised a series of short LM species (8-27 residues long) containing an α1-6-linked mannose backbone with greatly reduced α1-2-mannose side chains and no arabinose caps. The structure of the ΔNCgl2760 t-LM was similar to that of the t-LM produced by a C. glutamicum mutant lacking the mptA gene, encoding a membrane α1-6-mannosyltransferase involved in extending the α1-6-mannan backbone of LM intermediates. Interestingly, NCgl2760 lacks any motifs or homology to other proteins of known function. Attempts to delete the NCgl2760 orthologue in Mycobacterium smegmatis were unsuccessful, consistent with previous studies indicating that the M. tuberculosis orthologue, Rv0227c, is an essential gene. Together, these data suggest that NCgl2760/Rv0227c plays a critical role in the elongation of the mannan backbone of mycobacterial and corynebacterial LM, further highlighting the complexity of lipoglycan pathways of Corynebacterineae.
Keywords: Corynebacterium glutamicum; Mycobacterium tuberculosis; cell wall; glycolipid; lipid synthesis; lipoarabinomannan; lipomannan; membrane protein.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
Similar articles
-
Identification of a novel alpha(1-->6) mannopyranosyltransferase MptB from Corynebacterium glutamicum by deletion of a conserved gene, NCgl1505, affords a lipomannan- and lipoarabinomannan-deficient mutant.Mol Microbiol. 2008 Jun;68(6):1595-613. doi: 10.1111/j.1365-2958.2008.06265.x. Epub 2008 Apr 28. Mol Microbiol. 2008. PMID: 18452585 Free PMC article.
-
The lipoprotein LpqW is essential for the mannosylation of periplasmic glycolipids in Corynebacteria.J Biol Chem. 2012 Dec 14;287(51):42726-38. doi: 10.1074/jbc.M112.373415. Epub 2012 Oct 22. J Biol Chem. 2012. PMID: 23091062 Free PMC article.
-
Role of LmeA, a Mycobacterial Periplasmic Protein, in Maintaining the Mannosyltransferase MptA and Its Product Lipomannan under Stress.mSphere. 2020 Nov 4;5(6):e01039-20. doi: 10.1128/mSphere.01039-20. mSphere. 2020. PMID: 33148829 Free PMC article.
-
Mycobacterial lipoarabinomannan and related lipoglycans: from biogenesis to modulation of the immune response.Mol Microbiol. 2004 Jul;53(2):391-403. doi: 10.1111/j.1365-2958.2004.04183.x. Mol Microbiol. 2004. PMID: 15228522 Review.
-
Cell Wall Associated Factors of Mycobacterium tuberculosis as Major Virulence Determinants: Current Perspectives in Drugs Discovery and Design.Curr Drug Targets. 2017 Nov 30;18(16):1904-1918. doi: 10.2174/1389450118666170711150034. Curr Drug Targets. 2017. PMID: 28699515 Review.
Cited by
-
Structural basis for safe and efficient energy conversion in a respiratory supercomplex.Nat Commun. 2022 Jan 27;13(1):545. doi: 10.1038/s41467-022-28179-x. Nat Commun. 2022. PMID: 35087070 Free PMC article.
-
FhaA plays a key role in mycobacterial polar elongation and asymmetric growth.mBio. 2025 Mar 12;16(3):e0252624. doi: 10.1128/mbio.02526-24. Epub 2025 Jan 21. mBio. 2025. PMID: 39835815 Free PMC article.
-
MmpA, a Conserved Membrane Protein Required for Efficient Surface Transport of Trehalose Lipids in Corynebacterineae.Biomolecules. 2021 Nov 24;11(12):1760. doi: 10.3390/biom11121760. Biomolecules. 2021. PMID: 34944401 Free PMC article.
-
MtrP, a putative methyltransferase in Corynebacteria, is required for optimal membrane transport of trehalose mycolates.J Biol Chem. 2020 May 1;295(18):6108-6119. doi: 10.1074/jbc.RA119.011688. Epub 2020 Mar 26. J Biol Chem. 2020. PMID: 32217691 Free PMC article.
-
Adaptive laboratory evolution enhances methanol tolerance and conversion in engineered Corynebacterium glutamicum.Commun Biol. 2020 May 7;3(1):217. doi: 10.1038/s42003-020-0954-9. Commun Biol. 2020. PMID: 32382107 Free PMC article.
References
-
- Barry C. E. 3rd, and Mdluli K. (1996) Drug sensitivity and environmental adaptation of mycobacterial cell wall components. Trends Microbiol. 4, 275–281 - PubMed
-
- Jankute M., Cox J. A., Harrison J., and Besra G. S. (2015) Assembly of the mycobacterial cell wall. Annu. Rev. Microbiol. 69, 405–423 - PubMed
-
- Brennan P. J., and Nikaido H. (1995) The envelope of mycobacteria. Annu. Rev. Biochem. 64, 29–63 - PubMed
-
- Haites R. E., Morita Y. S., McConville M. J., and Billman-Jacobe H. (2005) Function of phosphatidylinositol in mycobacteria. J. Biol. Chem. 280, 10981–10987 - PubMed
-
- Skovierová H., Larrouy-Maumus G., Zhang J., Kaur D., Barilone N., Korduláková J., Gilleron M., Guadagnini S., Belanová M., Prevost M. C., Gicquel B., Puzo G., Chatterjee D., Brennan P. J., Nigou J., and Jackson M. (2009) AftD, a novel essential arabinofuranosyltransferase from mycobacteria. Glycobiology 19, 1235–1247 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
