

Intestinal NCoR1, a regulator of epithelial cell maturation, controls neonatal hyperbilirubinemia
- PMID: 28167773
- PMCID: PMC5338369
- DOI: 10.1073/pnas.1700232114
Intestinal NCoR1, a regulator of epithelial cell maturation, controls neonatal hyperbilirubinemia
Erratum in
-
Correction for Chen et al., Intestinal NCoR1, a regulator of epithelial cell maturation, controls neonatal hyperbilirubinemia.Proc Natl Acad Sci U S A. 2017 May 16;114(20):E4115. doi: 10.1073/pnas.1705671114. Epub 2017 May 8. Proc Natl Acad Sci U S A. 2017. PMID: 28484028 Free PMC article. No abstract available.
Abstract
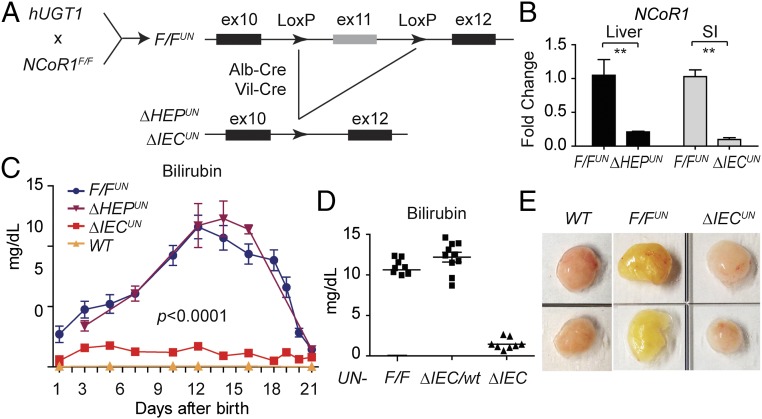
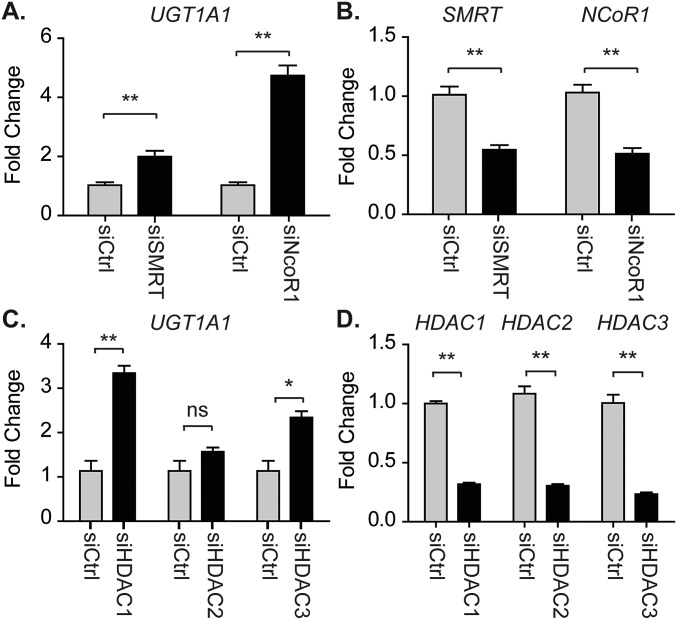
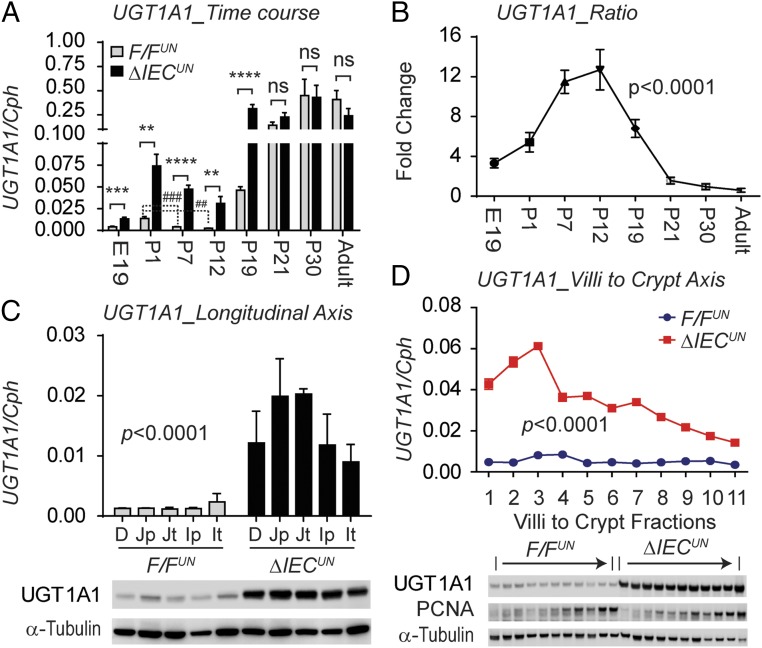
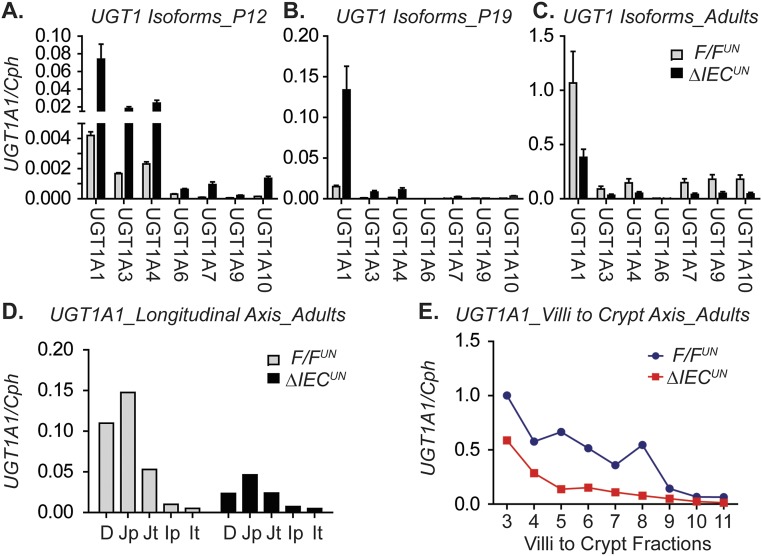
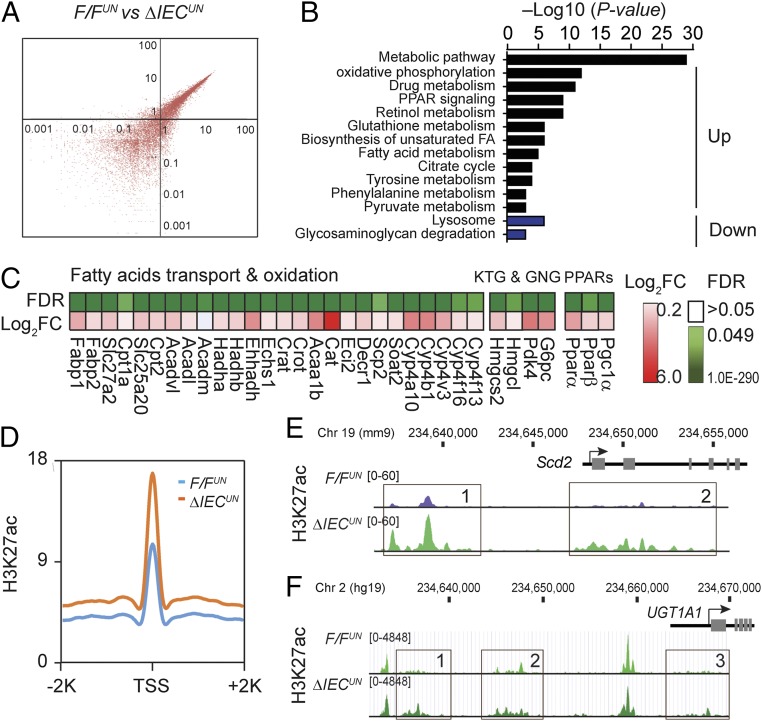
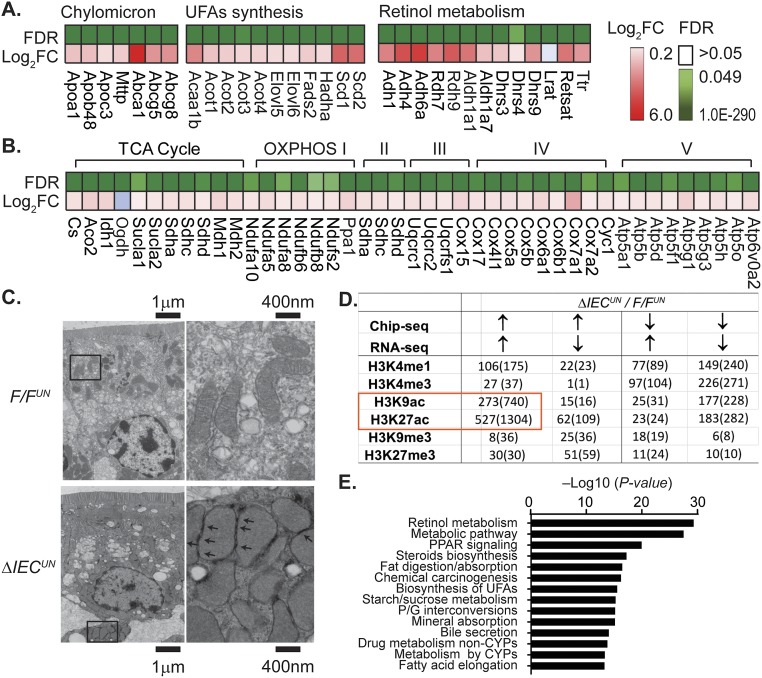
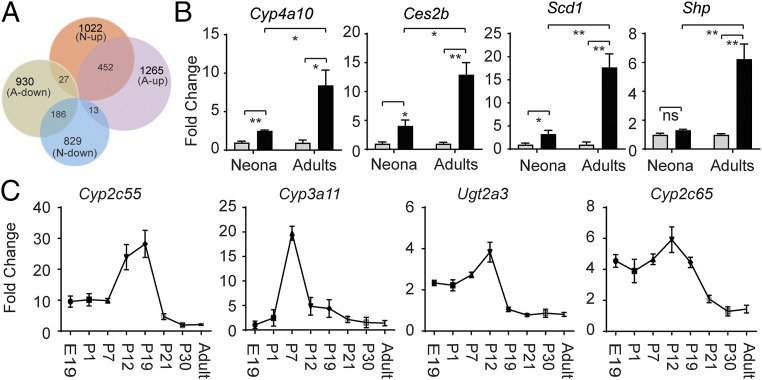
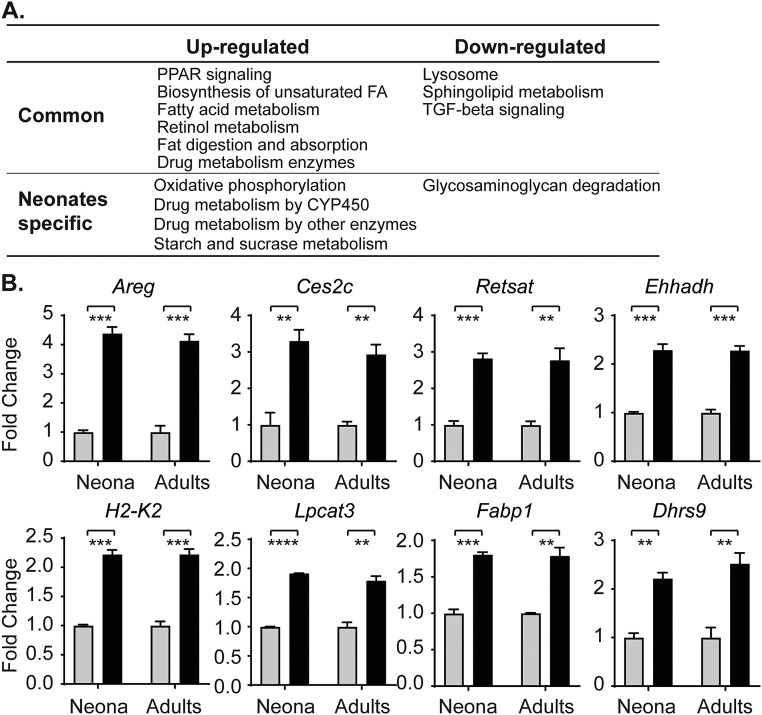
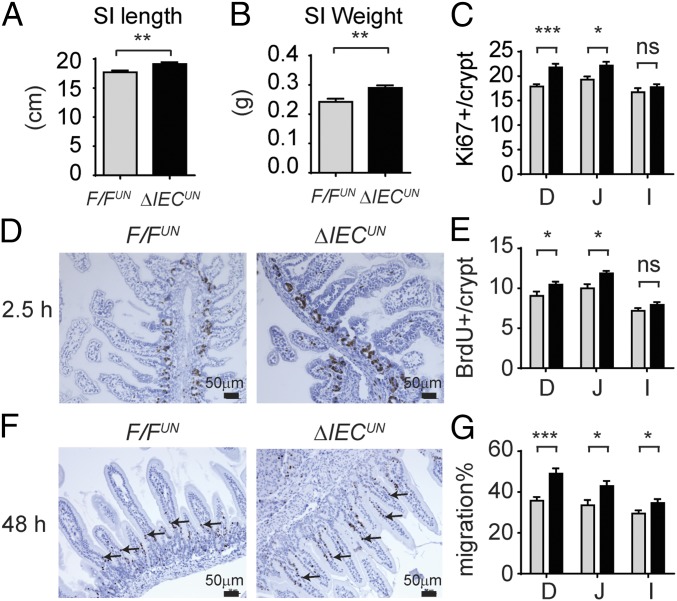
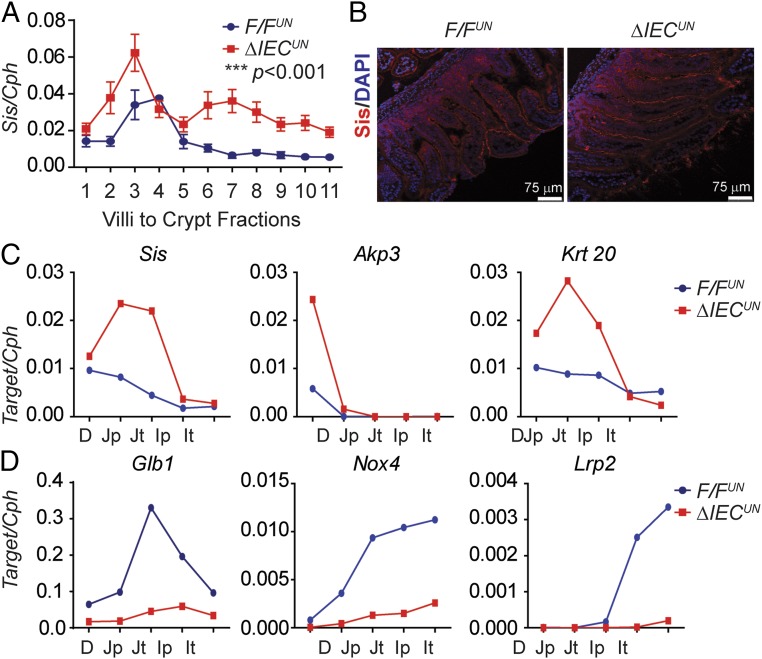
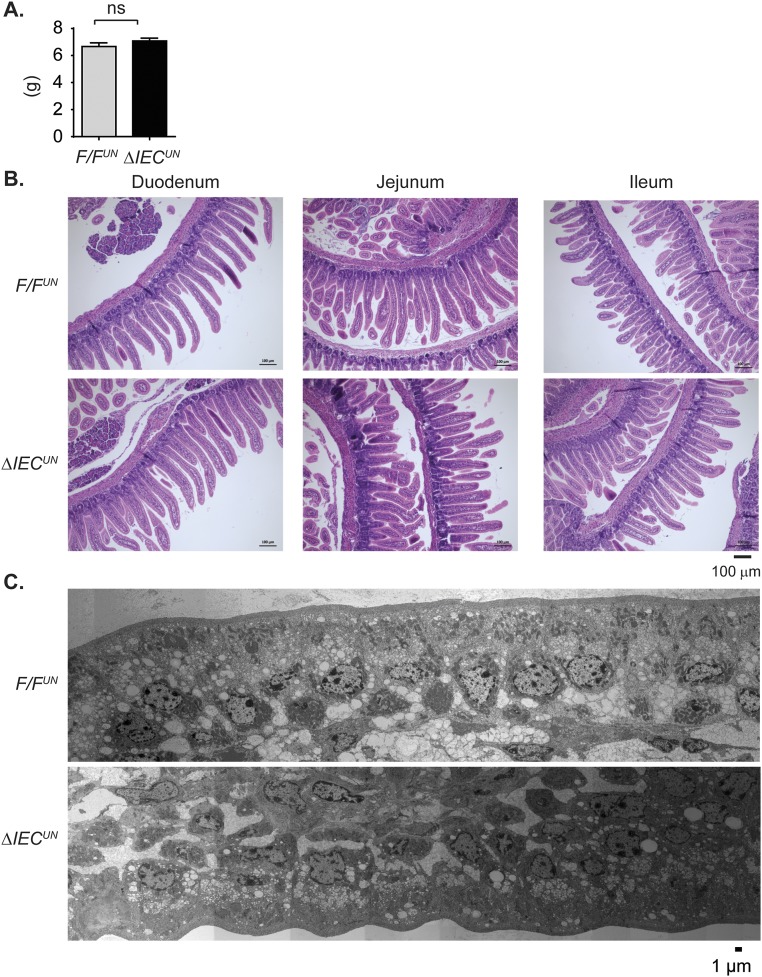
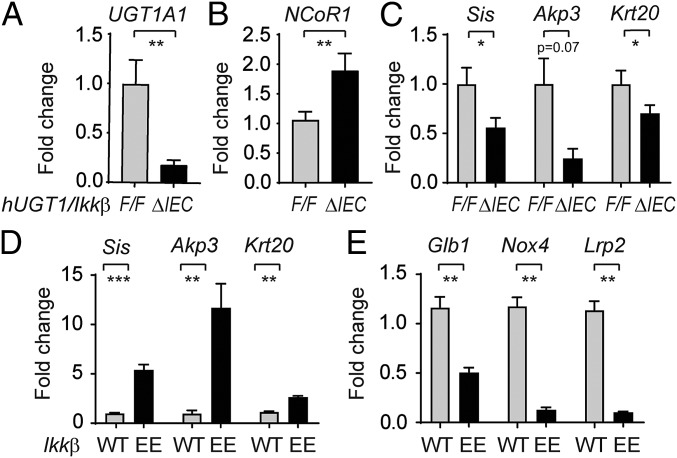
Severe neonatal hyperbilirubinemia (SNH) and the onset of bilirubin encephalopathy and kernicterus result in part from delayed expression of UDP-glucuronosyltransferase 1A1 (UGT1A1) and the inability to metabolize bilirubin. Although there is a good understanding of the early events after birth that lead to the rapid increase in serum bilirubin, the events that control delayed expression of UGT1A1 during development remain a mystery. Humanized UGT1 (hUGT1) mice develop SNH spontaneously, which is linked to repression of both liver and intestinal UGT1A1. In this study, we report that deletion of intestinal nuclear receptor corepressor 1 (NCoR1) completely diminishes hyperbilirubinemia in hUGT1 neonates because of intestinal UGT1A1 gene derepression. Transcriptomic studies and immunohistochemistry analysis demonstrate that NCoR1 plays a major role in repressing developmental maturation of the intestines. Derepression is marked by accelerated metabolic and oxidative phosphorylation, drug metabolism, fatty acid metabolism, and intestinal maturation, events that are controlled predominantly by H3K27 acetylation. The control of NCoR1 function and derepression is linked to IKKβ function, as validated in hUGT1 mice with targeted deletion of intestinal IKKβ. Physiological events during neonatal development that target activation of an IKKβ/NCoR1 loop in intestinal epithelial cells lead to derepression of genes involved in intestinal maturation and bilirubin detoxification. These findings provide a mechanism of NCoR1 in intestinal homeostasis during development and provide a key link to those events that control developmental repression of UGT1A1 and hyperbilirubinemia.
Keywords: IKKβ; UDP-glucuronosyltransferase 1A1; encephalopathy; humanized UGT1 mice; kernicterus.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Bosma PJ, et al. Bilirubin UDP-glucuronosyltransferase 1 is the only relevant bilirubin glucuronidating isoform in man. J Biol Chem. 1994;269(27):17960–17964. - PubMed
-
- Burchell B, et al. Development of human liver UDP-glucuronosyltransferases. Dev Pharmacol Ther. 1989;13(2-4):70–77. - PubMed
-
- Watchko JF. Identification of neonates at risk for hazardous hyperbilirubinemia: Emerging clinical insights. Pediatr Clin North Am. 2009;56(3):671–687. - PubMed
-
- Dennery PA, Seidman DS, Stevenson DK. Neonatal hyperbilirubinemia. N Engl J Med. 2001;344(8):581–590. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
