

Design principles of autocatalytic cycles constrain enzyme kinetics and force low substrate saturation at flux branch points
- PMID: 28169831
- PMCID: PMC5333975
- DOI: 10.7554/eLife.20667
Design principles of autocatalytic cycles constrain enzyme kinetics and force low substrate saturation at flux branch points
Abstract
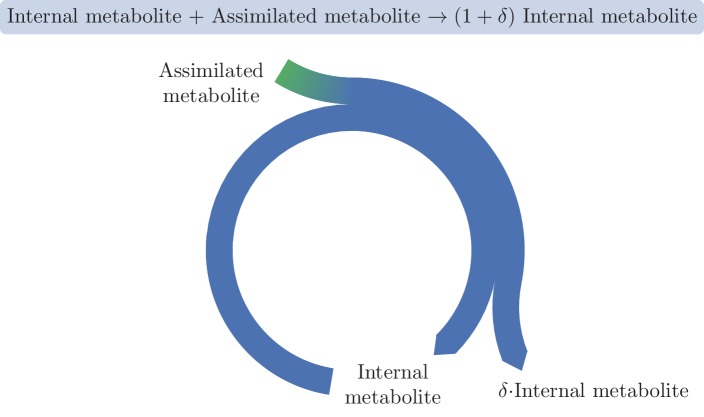
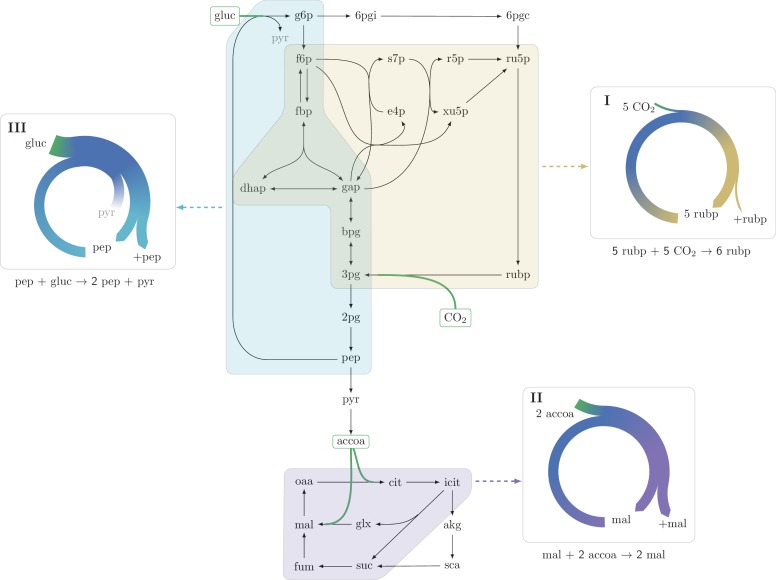
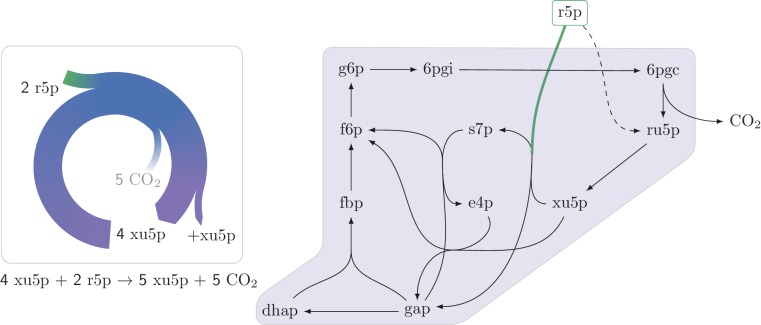
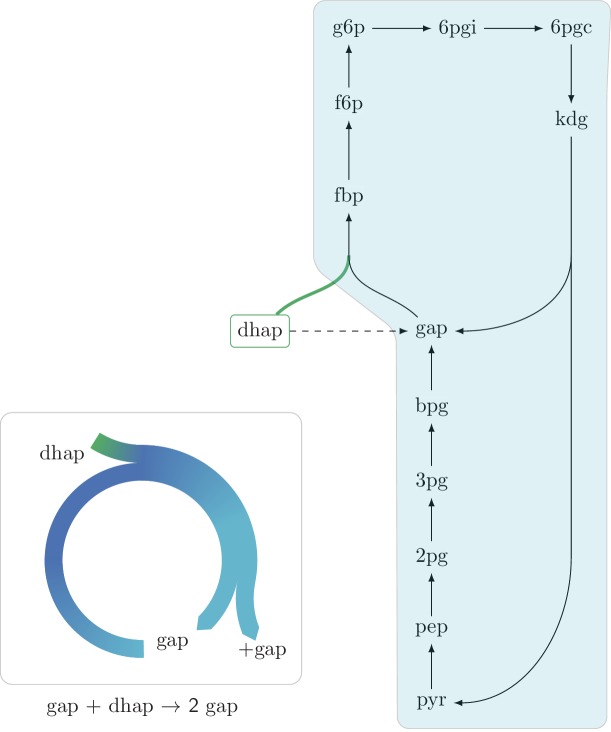
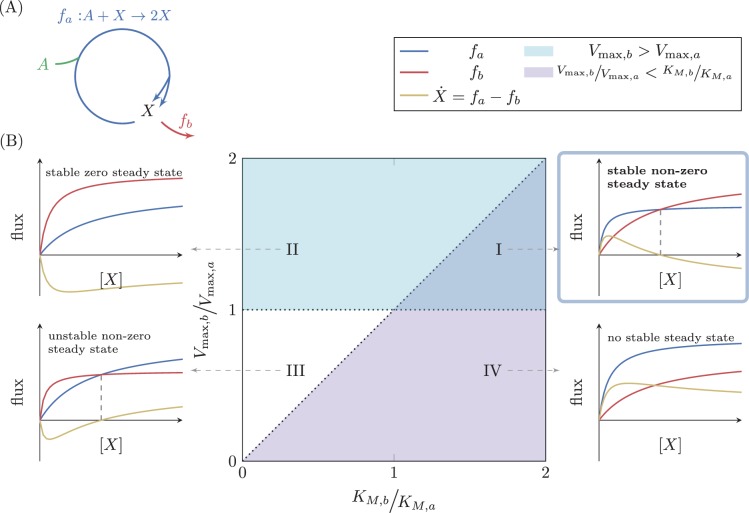
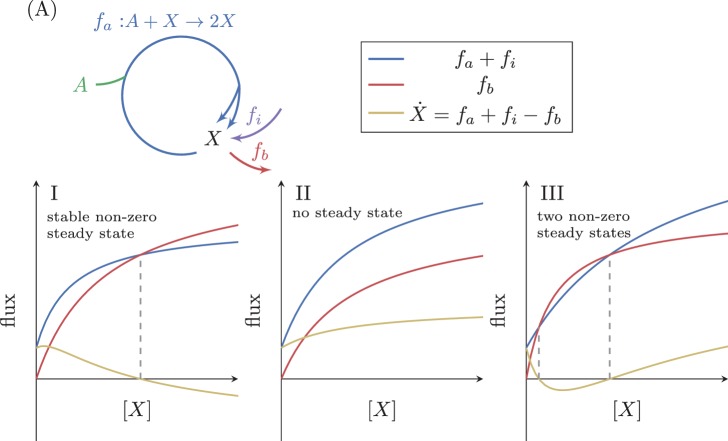
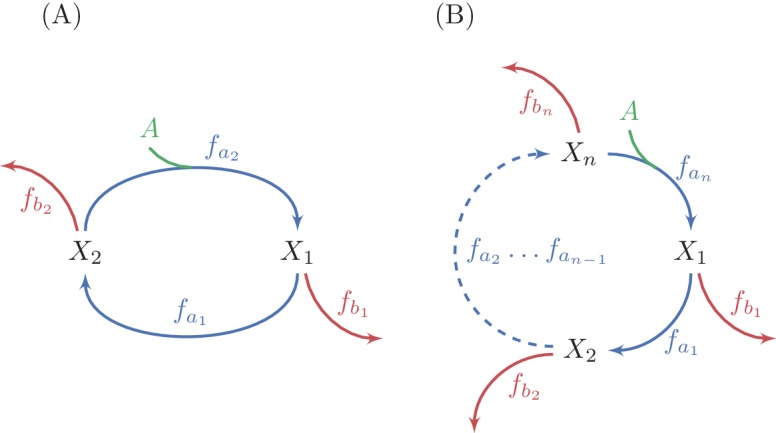
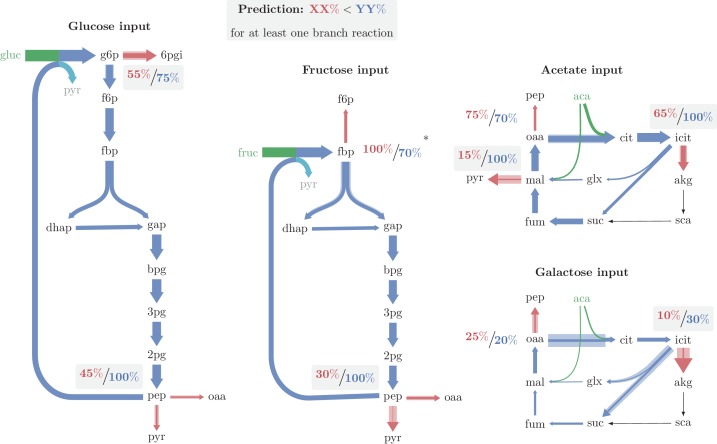
A set of chemical reactions that require a metabolite to synthesize more of that metabolite is an autocatalytic cycle. Here, we show that most of the reactions in the core of central carbon metabolism are part of compact autocatalytic cycles. Such metabolic designs must meet specific conditions to support stable fluxes, hence avoiding depletion of intermediate metabolites. As such, they are subjected to constraints that may seem counter-intuitive: the enzymes of branch reactions out of the cycle must be overexpressed and the affinity of these enzymes to their substrates must be relatively weak. We use recent quantitative proteomics and fluxomics measurements to show that the above conditions hold for functioning cycles in central carbon metabolism of E. coli. This work demonstrates that the topology of a metabolic network can shape kinetic parameters of enzymes and lead to seemingly wasteful enzyme usage.
Keywords: E. coli; computational biology; dynamic analysis; enzyme kinetics; metabolism; systems biology.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
References
-
- Antonovsky N, Gleizer S, Noor E, Zohar Y, Herz E, Barenholz U, Zelcbuch L, Amram S, Wides A, Tepper N, Davidi D, Bar-On Y, Bareia T, Wernick DG, Shani I, Malitsky S, Jona G, Bar-Even A, Milo R. Sugar synthesis from CO2 in Escherichia coli. Cell. 2016;166:115–125. doi: 10.1016/j.cell.2016.05.064. - DOI - PMC - PubMed
-
- Benson AA, Bassham JA, Calvin M, Goodale TC, Haas VA, Stepka W. The path of carbon in photosynthesis. v. paper chromatography and radioautography of the products1. Journal of the American Chemical Society. 1950;72:1710–1718. doi: 10.1021/ja01160a080. - DOI
-
- Davidi D, Noor E, Liebermeister W, Bar-Even A, Flamholz A, Tummler K, Barenholz U, Goldenfeld M, Shlomi T, Milo R. Global characterization of in vivo enzyme catalytic rates and their correspondence to in vitro kcat measurements. PNAS. 2016;113:3401–3406. doi: 10.1073/pnas.1514240113. - DOI - PMC - PubMed
-
- Eigen M, Schuster P. Springer Science & Business Media. 2012. The hypercycle: a principle of natural self-organization. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
