

The human RHOX gene cluster: target genes and functional analysis of gene variants in infertile men
- PMID: 28171660
- PMCID: PMC6281360
- DOI: 10.1093/hmg/ddw313
The human RHOX gene cluster: target genes and functional analysis of gene variants in infertile men
Abstract
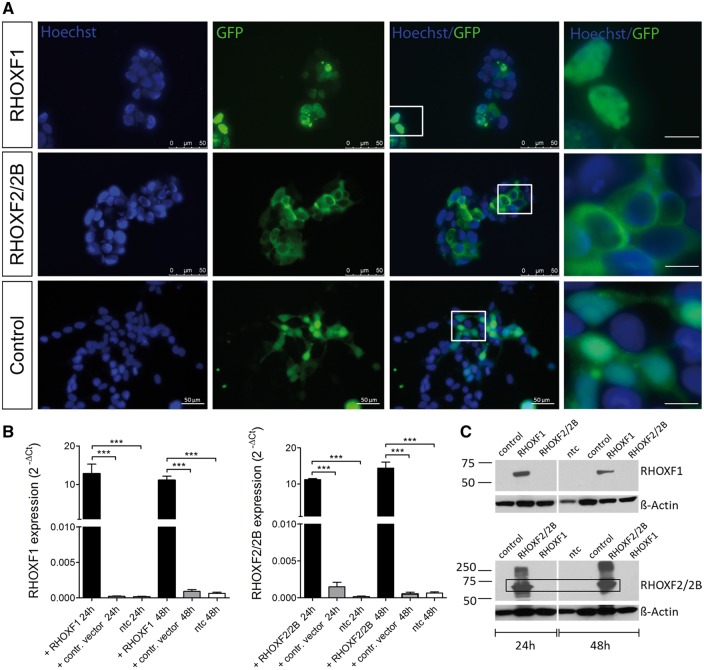
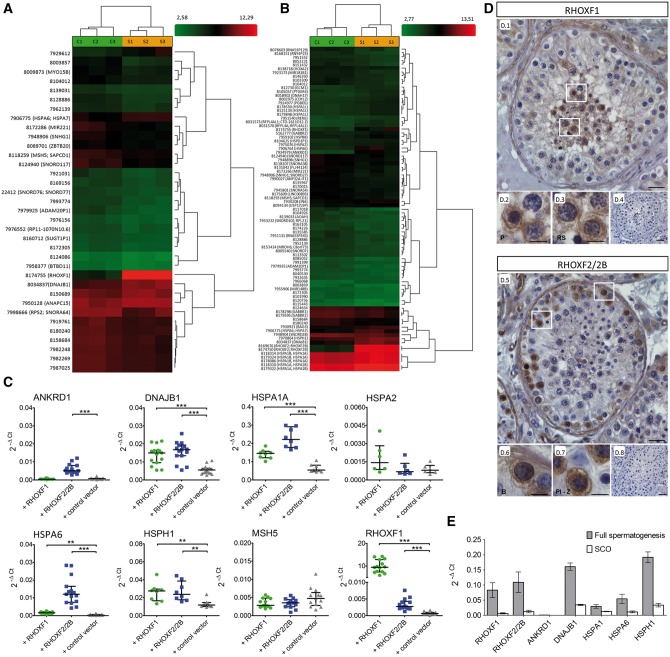
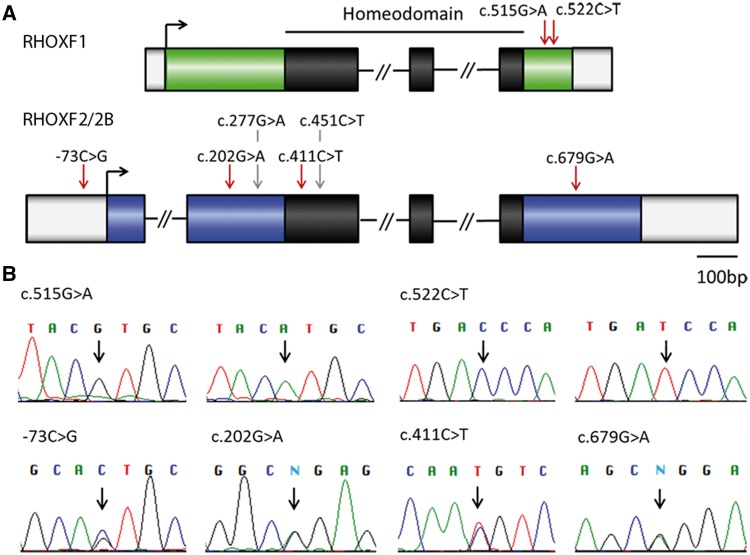
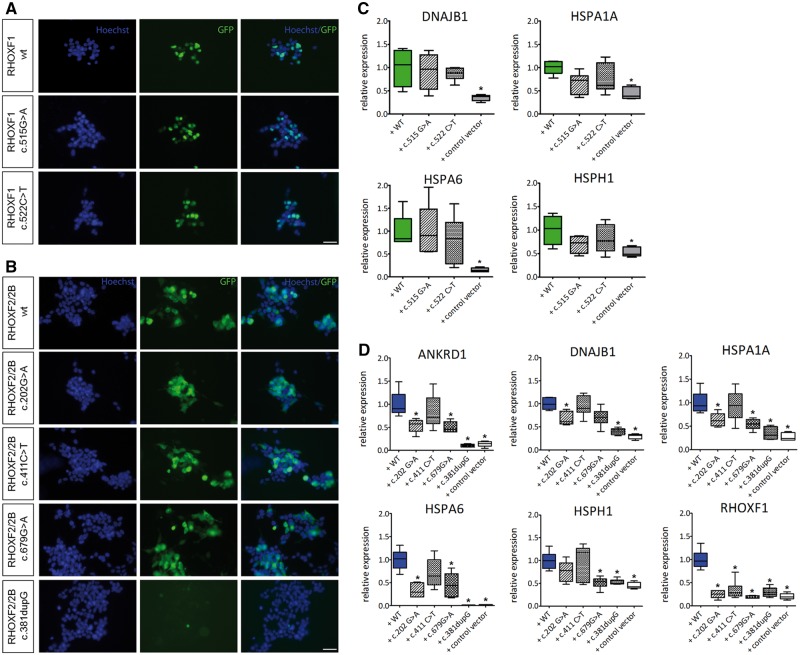
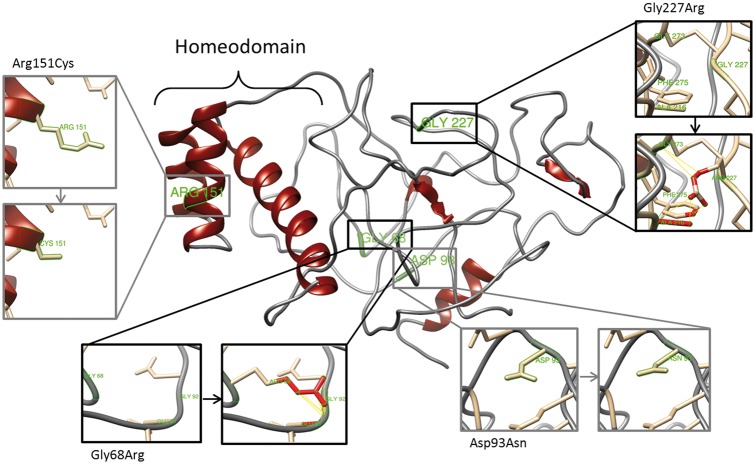
The X-linked reproductive homeobox (RHOX) gene cluster encodes transcription factors preferentially expressed in reproductive tissues. This gene cluster has important roles in male fertility based on phenotypic defects of Rhox-mutant mice and the finding that aberrant RHOX promoter methylation is strongly associated with abnormal human sperm parameters. However, little is known about the molecular mechanism of RHOX function in humans. Using gene expression profiling, we identified genes regulated by members of the human RHOX gene cluster. Some genes were uniquely regulated by RHOXF1 or RHOXF2/2B, while others were regulated by both of these transcription factors. Several of these regulated genes encode proteins involved in processes relevant to spermatogenesis; e.g. stress protection and cell survival. One of the target genes of RHOXF2/2B is RHOXF1, suggesting cross-regulation to enhance transcriptional responses. The potential role of RHOX in human infertility was addressed by sequencing all RHOX exons in a group of 250 patients with severe oligozoospermia. This revealed two mutations in RHOXF1 (c.515G > A and c.522C > T) and four in RHOXF2/2B (-73C > G, c.202G > A, c.411C > T and c.679G > A), of which only one (c.202G > A) was found in a control group of men with normal sperm concentration. Functional analysis demonstrated that c.202G > A and c.679G > A significantly impaired the ability of RHOXF2/2B to regulate downstream genes. Molecular modelling suggested that these mutations alter RHOXF2/F2B protein conformation. By combining clinical data with in vitro functional analysis, we demonstrate how the X-linked RHOX gene cluster may function in normal human spermatogenesis and we provide evidence that it is impaired in human male fertility.
Figures
References
-
- Tüttelmann F., Gromoll J., Kliesch S. (2008) Genetics of male infertility. Der. Urol., 47, 1561–1562. 1564–1567. - PubMed
-
- Tüttelmann F., Gromoll J. (2010) Novel genetic aspects of Klinefelter’s syndrome. Mol. Hum. Reprod., 16, 386–395. - PubMed
-
- Wang P.J., McCarrey J.R., Yang F., Page D.C. (2001) An abundance of X-linked genes expressed in spermatogonia. Nat. Genet., 27, 422–426. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
