

Nidovirus RNA polymerases: Complex enzymes handling exceptional RNA genomes
- PMID: 28174054
- PMCID: PMC7114556
- DOI: 10.1016/j.virusres.2017.01.023
Nidovirus RNA polymerases: Complex enzymes handling exceptional RNA genomes
Abstract
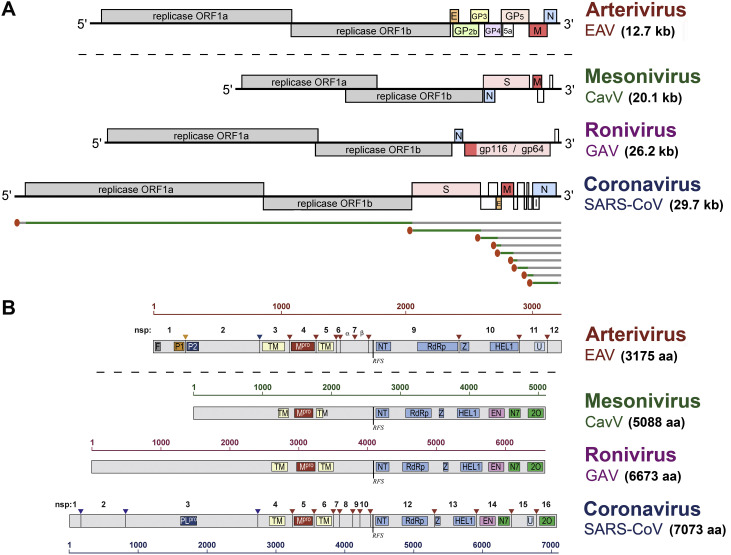
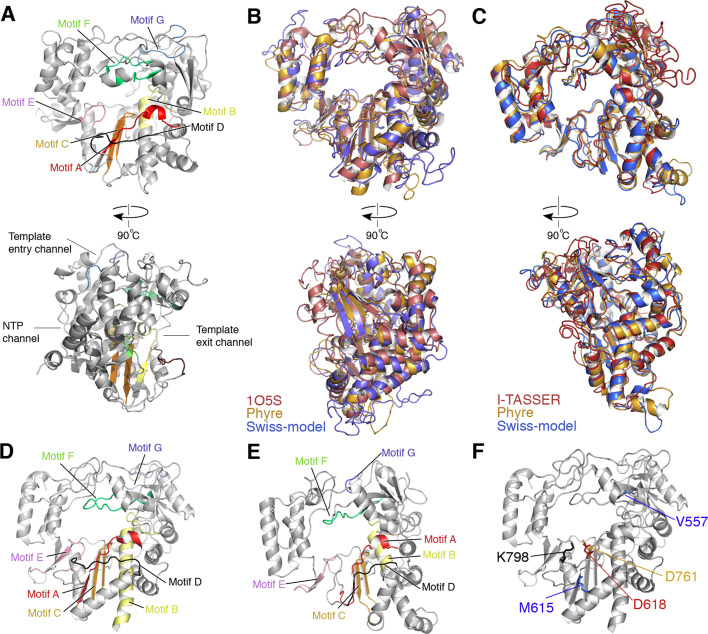
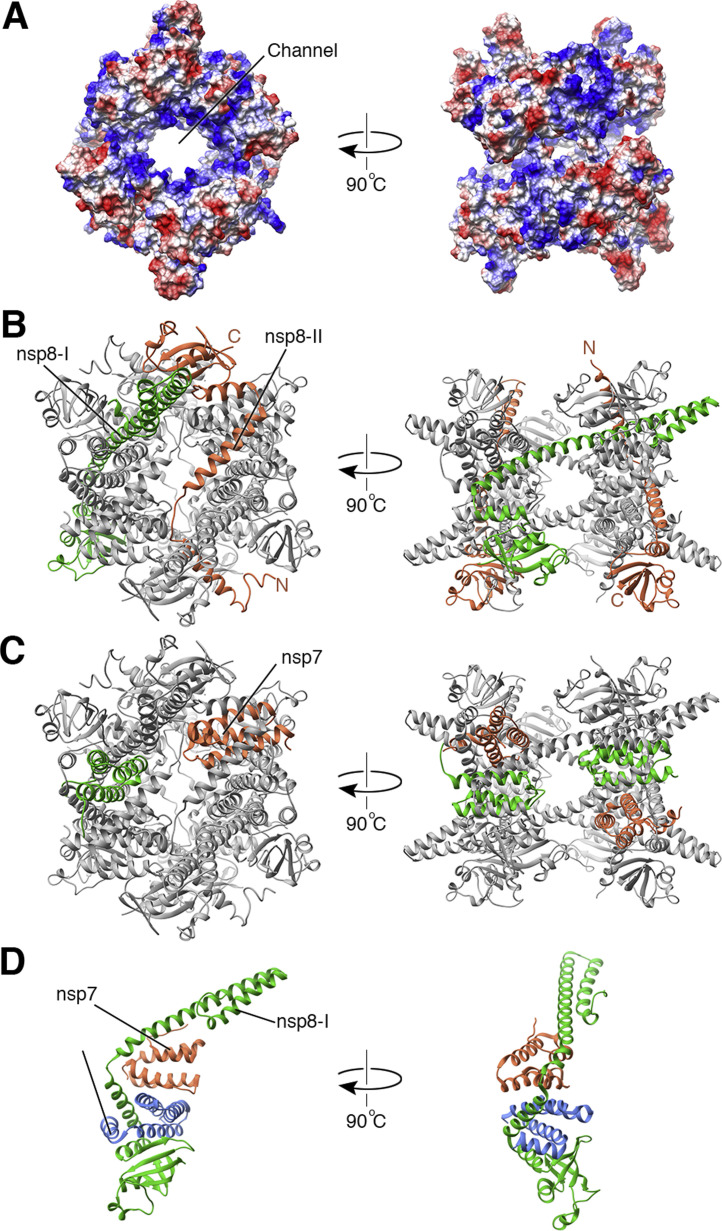
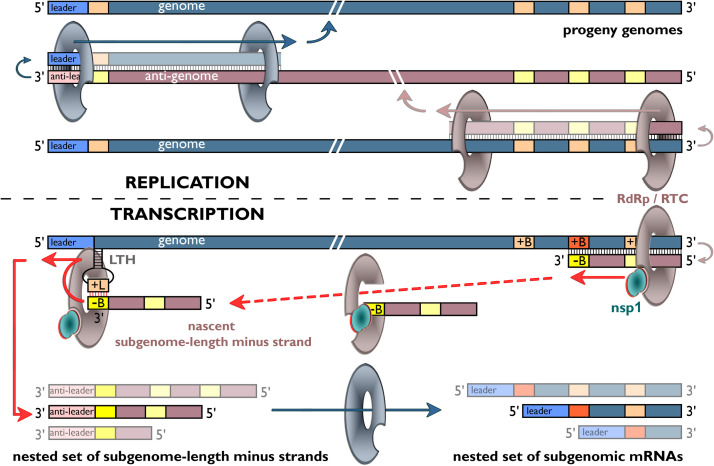
Coronaviruses and arteriviruses are distantly related human and animal pathogens that belong to the order Nidovirales. Nidoviruses are characterized by their polycistronic plus-stranded RNA genome, the production of subgenomic mRNAs and the conservation of a specific array of replicase domains, including key RNA-synthesizing enzymes. Coronaviruses (26-34 kilobases) have the largest known RNA genomes and their replication presumably requires a processive RNA-dependent RNA polymerase (RdRp) and enzymatic functions that suppress the consequences of the typically high error rate of viral RdRps. The arteriviruses have significantly smaller genomes and form an intriguing package with the coronaviruses to analyse viral RdRp evolution and function. The RdRp domain of nidoviruses resides in a cleavage product of the replicase polyprotein named non-structural protein (nsp) 12 in coronaviruses and nsp9 in arteriviruses. In all nidoviruses, the C-terminal RdRp domain is linked to a conserved N-terminal domain, which has been coined NiRAN (nidovirus RdRp-associated nucleotidyl transferase). Although no structural information is available, the functional characterization of the nidovirus RdRp and the larger enzyme complex of which it is part, has progressed significantly over the past decade. In coronaviruses several smaller, non-enzymatic nsps were characterized that direct RdRp function, while a 3'-to-5' exoribonuclease activity in nsp14 was implicated in fidelity. In arteriviruses, the nsp1 subunit was found to maintain the balance between genome replication and subgenomic mRNA production. Understanding RdRp behaviour and interactions during RNA synthesis and subsequent processing will be key to rationalising the evolutionary success of nidoviruses and the development of antiviral strategies.
Keywords: Arterivirus; Coronavirus; Polymerase fidelity; Processivity factors; Replication and transcription complex; Subgenomic mRNA synthesis.
Copyright © 2017 The Authors. Published by Elsevier B.V. All rights reserved.
Figures

References
-
- Banerjee D., Reddy K.R. Review article: safety and tolerability of direct-acting anti-viral agents in the new era of hepatitis C therapy. Aliment. Pharmacol. Ther. 2016;43:674–696. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
