

Patterned cell and matrix dynamics in branching morphogenesis
- PMID: 28174204
- PMCID: PMC5350520
- DOI: 10.1083/jcb.201610048
Patterned cell and matrix dynamics in branching morphogenesis
Abstract
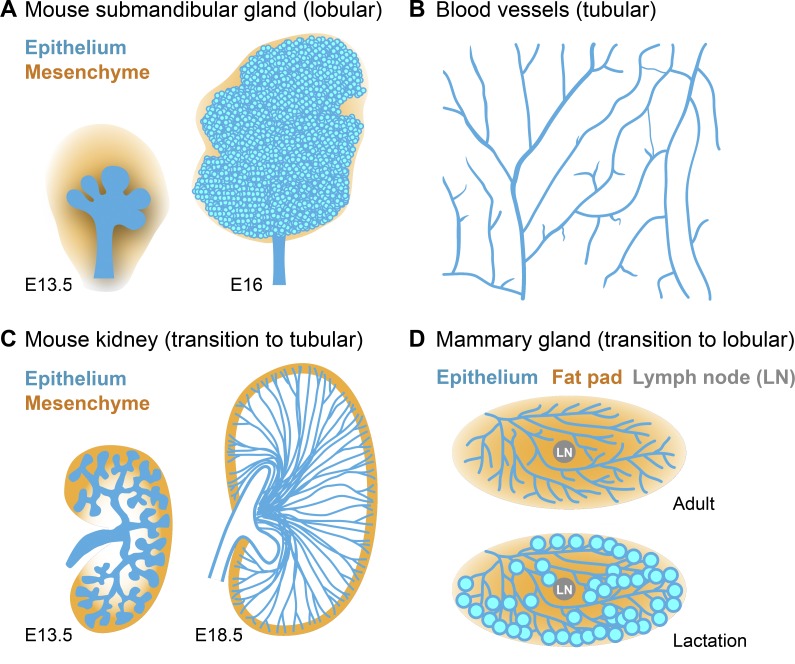
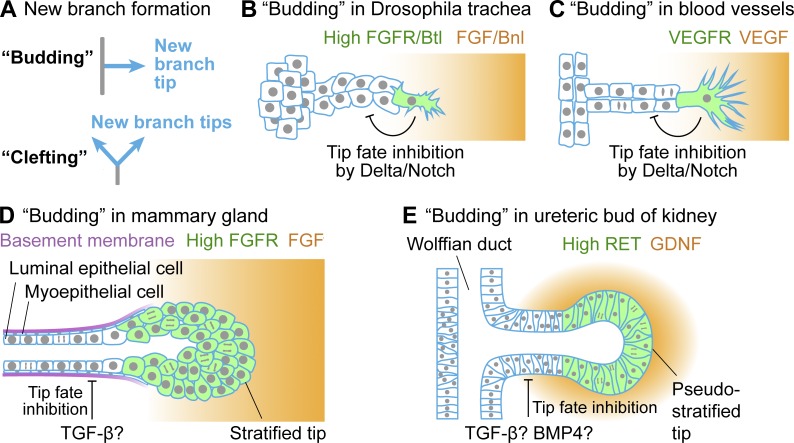
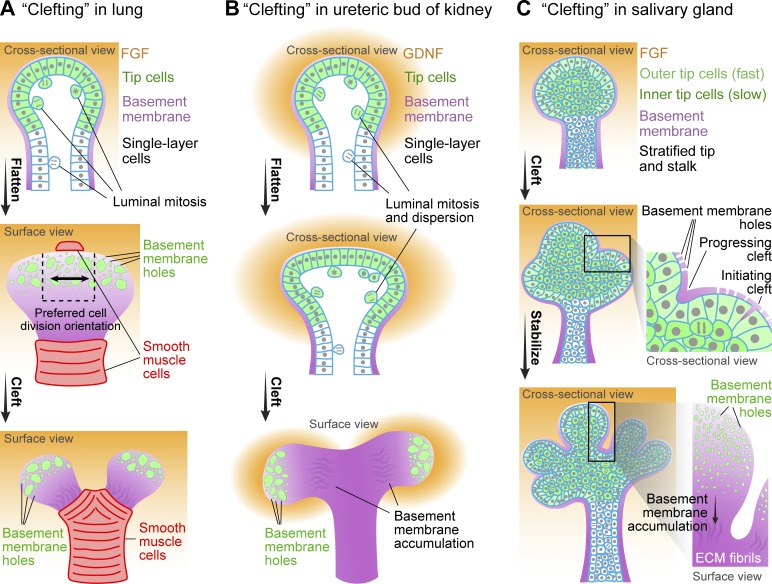
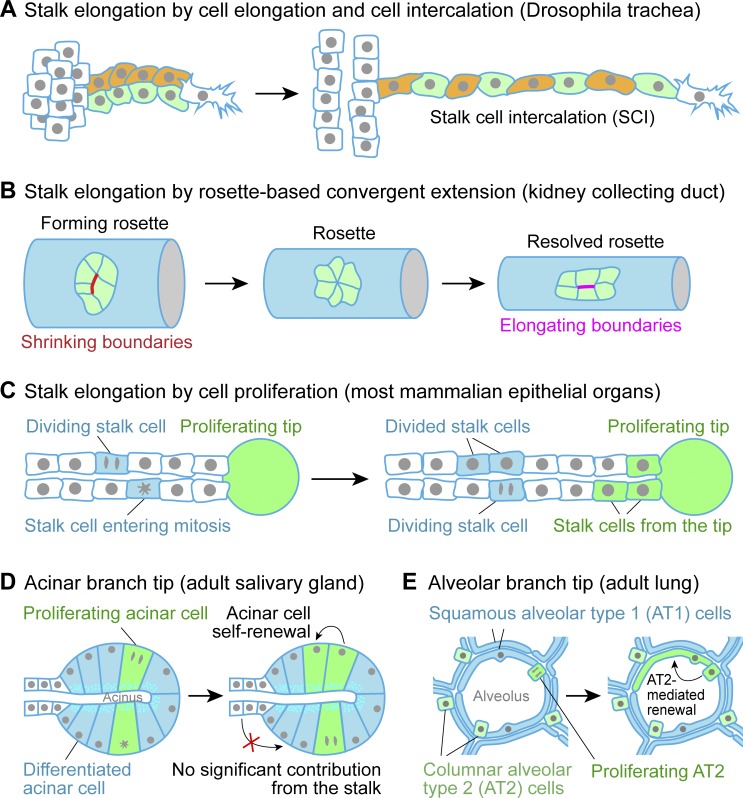
Many embryonic organs undergo branching morphogenesis to maximize their functional epithelial surface area. Branching morphogenesis requires the coordinated interplay of multiple types of cells with the extracellular matrix (ECM). During branching morphogenesis, new branches form by "budding" or "clefting." Cell migration, proliferation, rearrangement, deformation, and ECM dynamics have varied roles in driving budding versus clefting in different organs. Elongation of the newly formed branch and final maturation of the tip involve cellular mechanisms that include cell elongation, intercalation, convergent extension, proliferation, and differentiation. New methodologies such as high-resolution live imaging, tension sensors, and force-mapping techniques are providing exciting new opportunities for future research into branching morphogenesis.
This is a work of the U.S. Government and is not subject to copyright protection in the United States. Foreign copyrights may apply.
Figures
References
-
- Akhtar N., Marlow R., Lambert E., Schatzmann F., Lowe E.T., Cheung J., Katz E., Li W., Wu C., Dedhar S., et al. . 2009. Molecular dissection of integrin signalling proteins in the control of mammary epithelial development and differentiation. Development. 136:1019–1027. 10.1242/dev.028423 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
