

Central Regulatory Role for SIN1 in Interferon γ (IFNγ) Signaling and Generation of Biological Responses
- PMID: 28174303
- PMCID: PMC5377787
- DOI: 10.1074/jbc.M116.757666
Central Regulatory Role for SIN1 in Interferon γ (IFNγ) Signaling and Generation of Biological Responses
Abstract
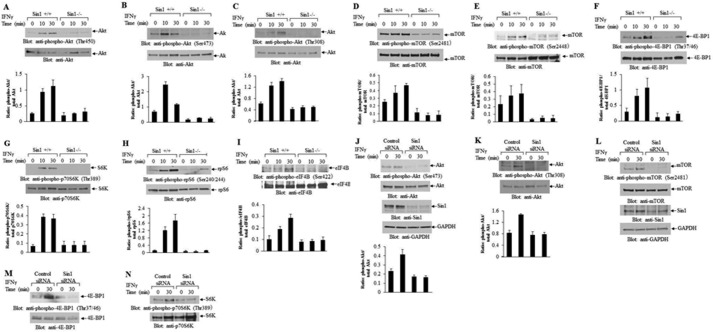
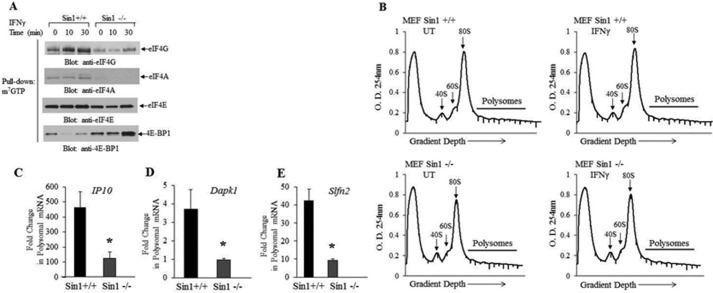
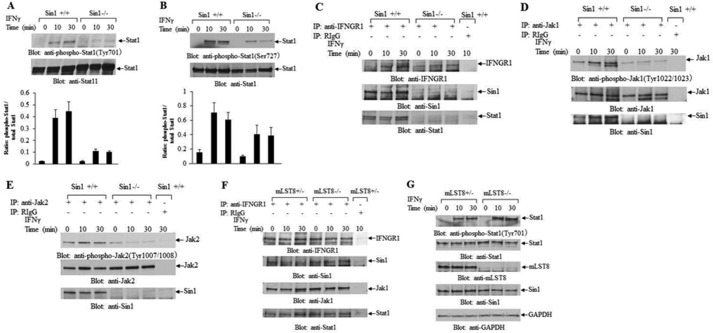
The precise signaling mechanisms by which type II IFN receptors control expression of unique genes to induce biological responses remain to be established. We provide evidence that Sin1, a known element of the mammalian target of rapamycin complex 2 (mTORC2), is required for IFNγ-induced phosphorylation and activation of AKT and that such activation mediates downstream regulation of mTORC1 and its effectors. These events play important roles in the assembly of the eukaryotic translation initiation factor 4F (eIF4F) and mRNA translation of IFN-stimulated genes. Interestingly, IFNγ-induced tyrosine phosphorylation of STAT1 is reduced in cells with targeted disruption of Sin1, leading to decreased transcription of several IFNγ-inducible genes in an mTORC2-independent manner. Additionally, our studies establish that Sin1 is essential for generation of type II IFN-dependent antiviral effects and antiproliferative responses in normal and malignant hematopoiesis. Together, our findings establish an important role for Sin1 in both transcription and translation of IFN-stimulated genes and type II IFN-mediated biological responses, involving both mTORC2-dependent and -independent functions.
Keywords: RNA editing; antiviral agent; innate immunity; interferon; signal transduction; translation control.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures




Similar articles
-
Interferon γ (IFNγ) Signaling via Mechanistic Target of Rapamycin Complex 2 (mTORC2) and Regulatory Effects in the Generation of Type II Interferon Biological Responses.J Biol Chem. 2016 Jan 29;291(5):2389-96. doi: 10.1074/jbc.M115.664995. Epub 2015 Dec 8. J Biol Chem. 2016. PMID: 26645692 Free PMC article.
-
Sin1 (Stress-Activated Protein Kinase-Interacting Protein) Regulates Ischemia-Induced Microthrombosis Through Integrin αIIbβ3-Mediated Outside-In Signaling and Hypoxia Responses in Platelets.Arterioscler Thromb Vasc Biol. 2018 Dec;38(12):2793-2805. doi: 10.1161/ATVBAHA.118.311822. Arterioscler Thromb Vasc Biol. 2018. PMID: 30571167
-
Critical roles for Rictor/Sin1 complexes in interferon-dependent gene transcription and generation of antiproliferative responses.J Biol Chem. 2014 Mar 7;289(10):6581-6591. doi: 10.1074/jbc.M113.537852. Epub 2014 Jan 27. J Biol Chem. 2014. PMID: 24469448 Free PMC article.
-
STAT1 as a novel therapeutical target in pro-atherogenic signal integration of IFNγ, TLR4 and IL-6 in vascular disease.Cytokine Growth Factor Rev. 2011 Aug;22(4):211-9. doi: 10.1016/j.cytogfr.2011.06.003. Epub 2011 Jul 12. Cytokine Growth Factor Rev. 2011. PMID: 21752694 Review.
-
Akt and mRNA translation by interferons.Cell Cycle. 2008 Jul 15;7(14):2112-6. doi: 10.4161/cc.7.14.6258. Epub 2008 May 8. Cell Cycle. 2008. PMID: 18635959 Review.
Cited by
-
Unmasking the tumourigenic role of SIN1/MAPKAP1 in the mTOR complex 2.Clin Transl Med. 2023 Oct;13(10):e1464. doi: 10.1002/ctm2.1464. Clin Transl Med. 2023. PMID: 37877351 Free PMC article. Review.
-
Folliculin regulates mTORC1/2 and WNT pathways in early human pluripotency.Nat Commun. 2019 Feb 7;10(1):632. doi: 10.1038/s41467-018-08020-0. Nat Commun. 2019. PMID: 30733432 Free PMC article.
-
IFN-γ-inducible antiviral responses require ULK1-mediated activation of MLK3 and ERK5.Sci Signal. 2018 Nov 20;11(557):eaap9921. doi: 10.1126/scisignal.aap9921. Sci Signal. 2018. PMID: 30459284 Free PMC article.
-
Sin1-mediated mTOR signaling in cell growth, metabolism and immune response.Natl Sci Rev. 2019 Nov;6(6):1149-1162. doi: 10.1093/nsr/nwz171. Epub 2019 Nov 19. Natl Sci Rev. 2019. PMID: 34691993 Free PMC article.
-
TCDD-mediated suppression of naïve human B cell IgM secretion involves aryl hydrocarbon receptor-mediated reduction in STAT3 serine 727 phosphorylation and is restored by interferon-γ.Cell Signal. 2020 Jan;65:109447. doi: 10.1016/j.cellsig.2019.109447. Epub 2019 Oct 31. Cell Signal. 2020. PMID: 31678681 Free PMC article.
References
-
- Stark G. R., Kerr I. M., Williams B. R., Silverman R. H., and Schreiber R. D. (1998) How cells respond to interferons. Annu. Rev. Biochem. 67, 227–264 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
