

Foxn1-β5t transcriptional axis controls CD8+ T-cell production in the thymus
- PMID: 28176764
- PMCID: PMC5309848
- DOI: 10.1038/ncomms14419
Foxn1-β5t transcriptional axis controls CD8+ T-cell production in the thymus
Abstract
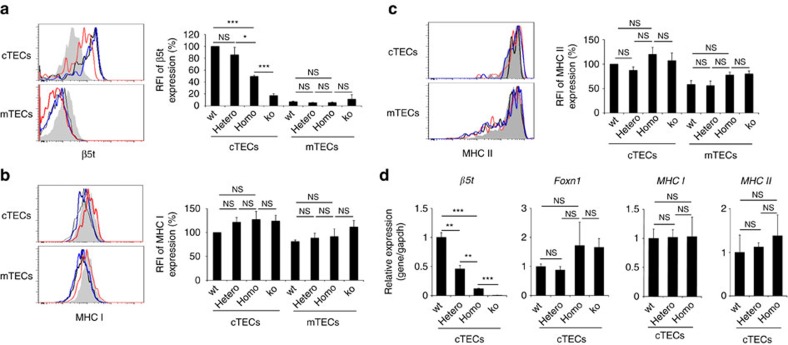
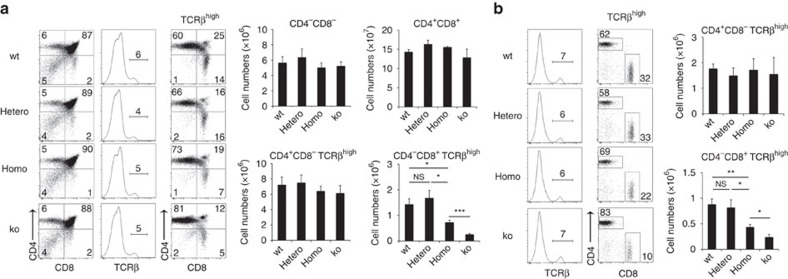
The thymus is an organ that produces functionally competent T cells that protect us from pathogens and malignancies. Foxn1 is a transcription factor that is essential for thymus organogenesis; however, the direct target for Foxn1 to actuate thymic T-cell production is unknown. Here we show that a Foxn1-binding cis-regulatory element promotes the transcription of β5t, which has an essential role in cortical thymic epithelial cells to induce positive selection of functionally competent CD8+ T cells. A point mutation in this genome element results in a defect in β5t expression and CD8+ T-cell production in mice. The results reveal a Foxn1-β5t transcriptional axis that governs CD8+ T-cell production in the thymus.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Doherty P. C. & Christensen J. P. Accessing complexity: the dynamics of virus-specific T cell responses. Annu. Rev. Immunol. 18, 561–592 (2000). - PubMed
-
- Wong P. & Pamer E. G. CD8 T cell responses to infectious pathogens. Annu. Rev. Immunol. 21, 29–70 (2003). - PubMed
-
- Appay V., Douek D. C. & Price D. A. CD8+ T cell efficacy in vaccination and disease. Nat. Med. 14, 623–628 (2008). - PubMed
-
- Fink P. J. & Bevan M. J. Positive selection of thymocytes. Adv. Immunol. 59, 99–133 (1995). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
