

Structural Insights into the Broad Substrate Specificity of a Novel Endoglycoceramidase I Belonging to a New Subfamily of GH5 Glycosidases
- PMID: 28179425
- PMCID: PMC5377795
- DOI: 10.1074/jbc.M116.763821
Structural Insights into the Broad Substrate Specificity of a Novel Endoglycoceramidase I Belonging to a New Subfamily of GH5 Glycosidases
Abstract
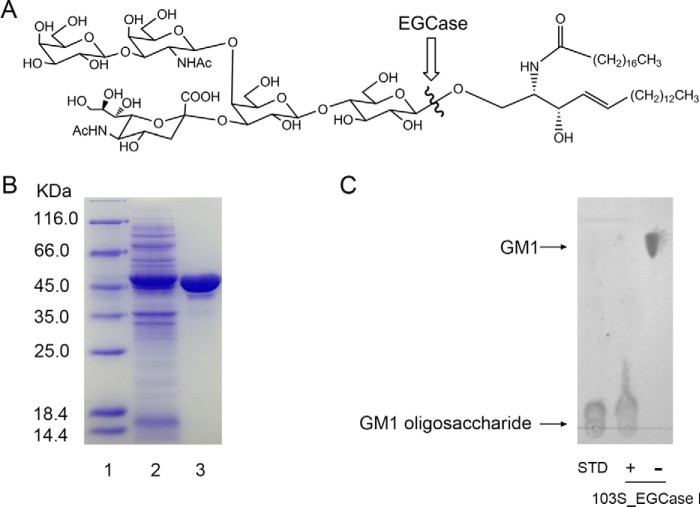
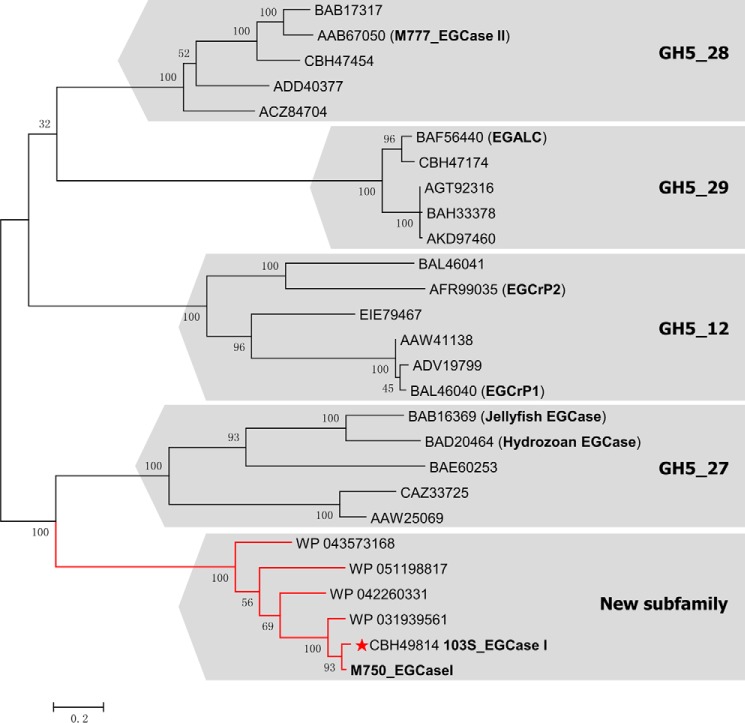
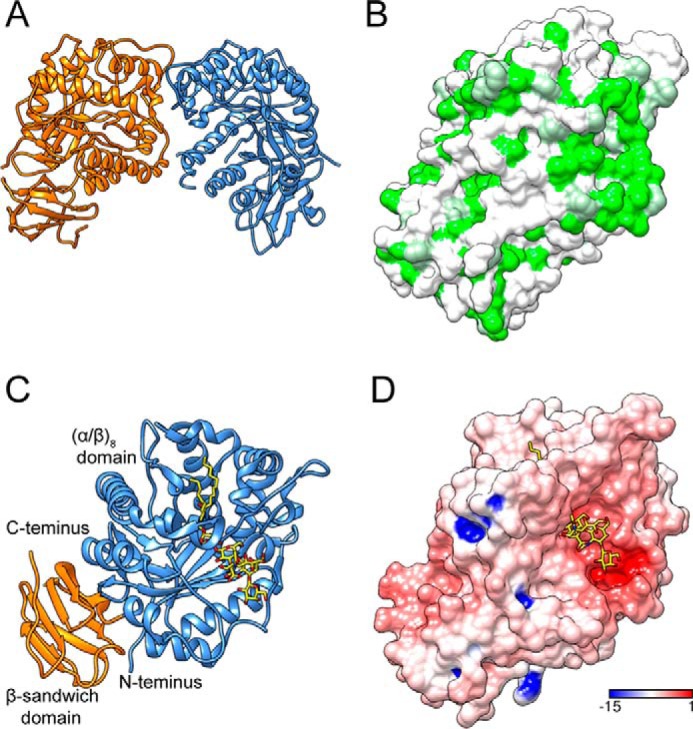
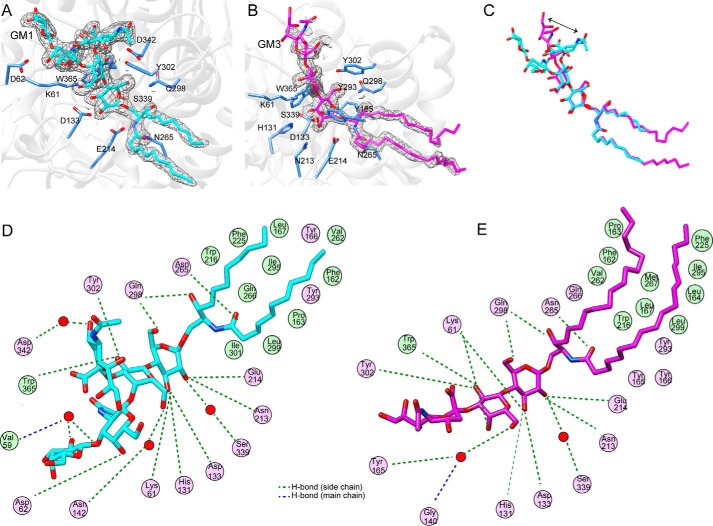
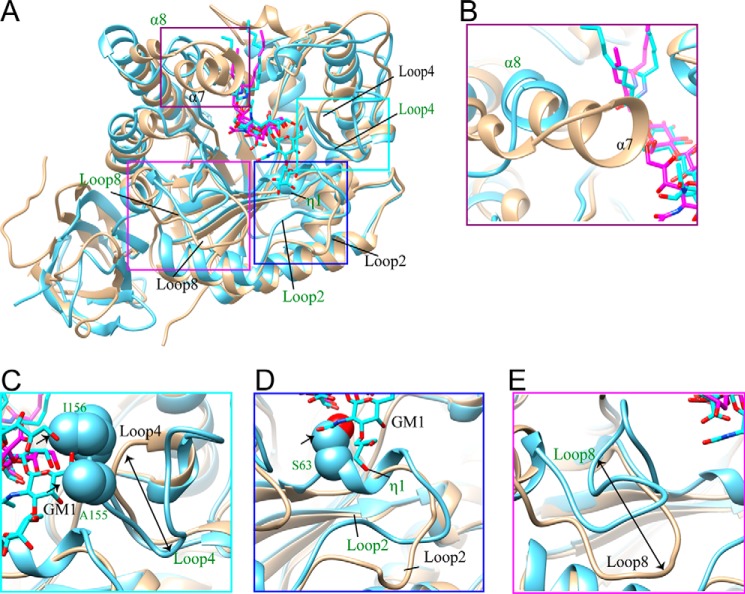
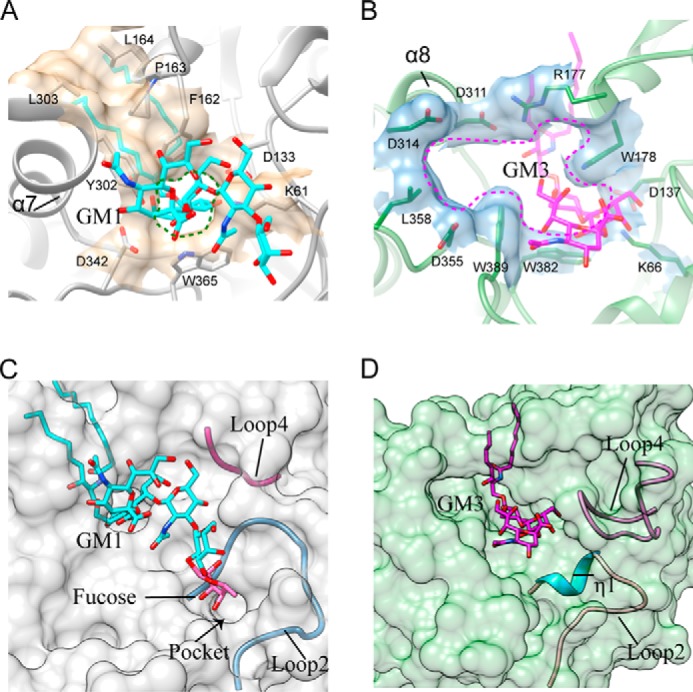
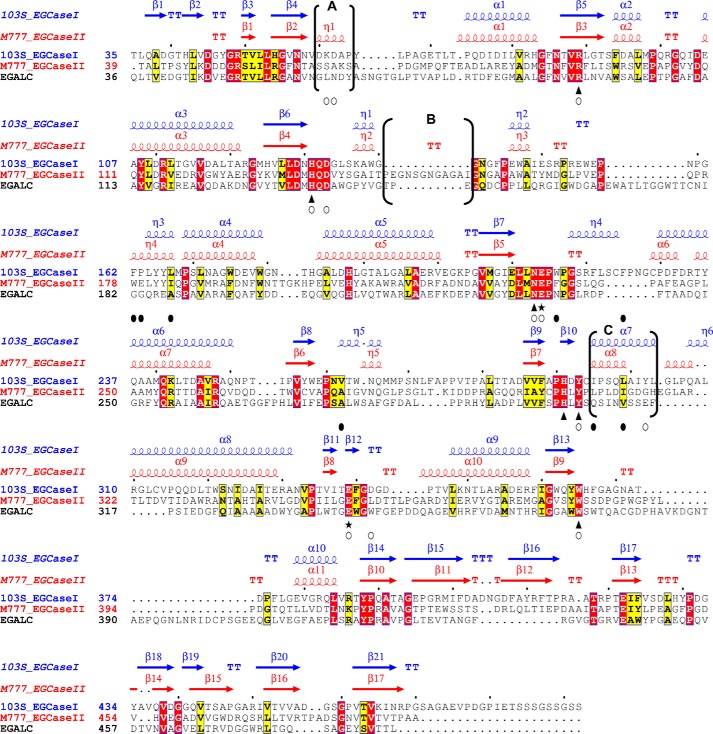
Endoglycoceramidases (EGCases) specifically hydrolyze the glycosidic linkage between the oligosaccharide and the ceramide moieties of various glycosphingolipids, and they have received substantial attention in the emerging field of glycosphingolipidology. However, the mechanism regulating the strict substrate specificity of these GH5 glycosidases has not been identified. In this study, we report a novel EGCase I from Rhodococcus equi 103S (103S_EGCase I) with remarkably broad substrate specificity. Based on phylogenetic analyses, the enzyme may represent a new subfamily of GH5 glycosidases. The X-ray crystal structures of 103S_EGCase I alone and in complex with its substrates monosialodihexosylganglioside (GM3) and monosialotetrahexosylganglioside (GM1) enabled us to identify several structural features that may account for its broad specificity. Compared with EGCase II from Rhodococcus sp. M-777 (M777_EGCase II), which possesses strict substrate specificity, 103S_EGCase I possesses a longer α7-helix and a shorter loop 4, which forms a larger substrate-binding pocket that could accommodate more extended oligosaccharides. In addition, loop 2 and loop 8 of the enzyme adopt a more open conformation, which also enlarges the oligosaccharide-binding cavity. Based on this knowledge, a rationally designed experiment was performed to examine the substrate specificity of EGCase II. The truncation of loop 4 in M777_EGCase II increased its activity toward GM1 (163%). Remarkably, the S63G mutant of M777_EGCase II showed a broader substrate spectra and significantly increased activity toward bulky substrates (up to >1370-fold for fucosyl-GM1). Collectively, the results presented here reveal the exquisite substrate recognition mechanism of EGCases and provide an opportunity for further engineering of these enzymes.
Keywords: crystal structure; glycoside hydrolase; protein engineering; sphingolipid; substrate specificity.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Fujitani N., Takegawa Y., Ishibashi Y., Araki K., Furukawa J., Mitsutake S., Igarashi Y., Ito M., and Shinohara Y. (2011) Qualitative and quantitative cellular glycomics of glycosphingolipids based on rhodococcal endoglycosylceramidase-assisted glycan cleavage, glycoblotting-assisted sample preparation, and matrix-assisted laser desorption ionization tandem time-of-flight mass spectrometry analysis. J. Biol. Chem. 286, 41669–41679 - PMC - PubMed
-
- Ito M., and Yamagata T. (1986) A novel glycosphingolipid-degrading enzyme cleaves the linkage between the oligosaccharide and ceramide of neutral and acidic glycosphingolipids. J. Biol. Chem. 261, 14278–14282 - PubMed
-
- Ashida H., Yamamoto K., Kumagai H., and Tochikura T. (1992) Purification and characterization of membrane-bound endoglycoceramidase from Corynebacterium sp. Eur. J. Biochem. 205, 729–735 - PubMed
-
- Li S.-C., DeGasperi R., Muldrey J. E., and Li Y.-T. (1986) A unique glycosphingolipid-splitting enzyme (ceramide-glycanase from leech) cleaves the linkage between the oligosaccharide and the ceramide. Biochem. Biophys. Res. Commun. 141, 346–352 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
