

Asparagine-Linked Glycans of Cryptosporidium parvum Contain a Single Long Arm, Are Barely Processed in the Endoplasmic Reticulum (ER) or Golgi, and Show a Strong Bias for Sites with Threonine
- PMID: 28179475
- PMCID: PMC5393390
- DOI: 10.1074/mcp.M116.066035
Asparagine-Linked Glycans of Cryptosporidium parvum Contain a Single Long Arm, Are Barely Processed in the Endoplasmic Reticulum (ER) or Golgi, and Show a Strong Bias for Sites with Threonine
Abstract
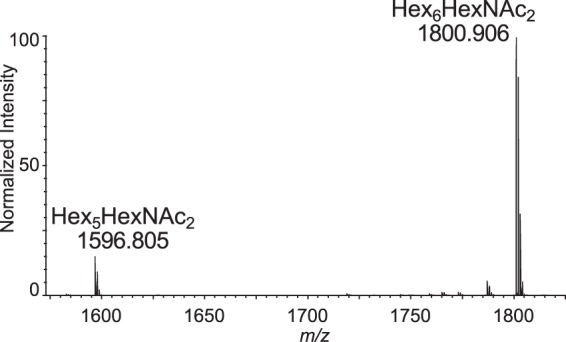
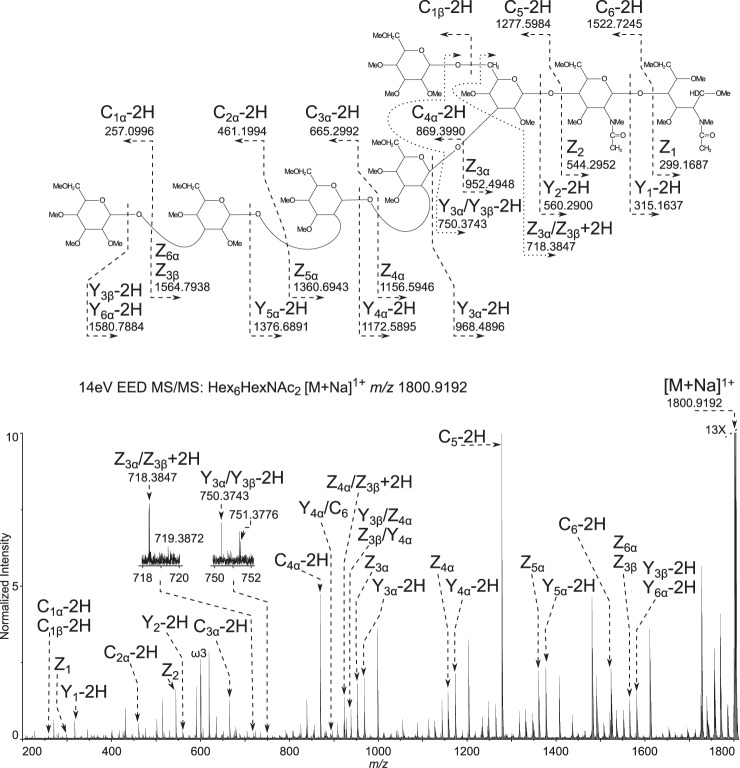
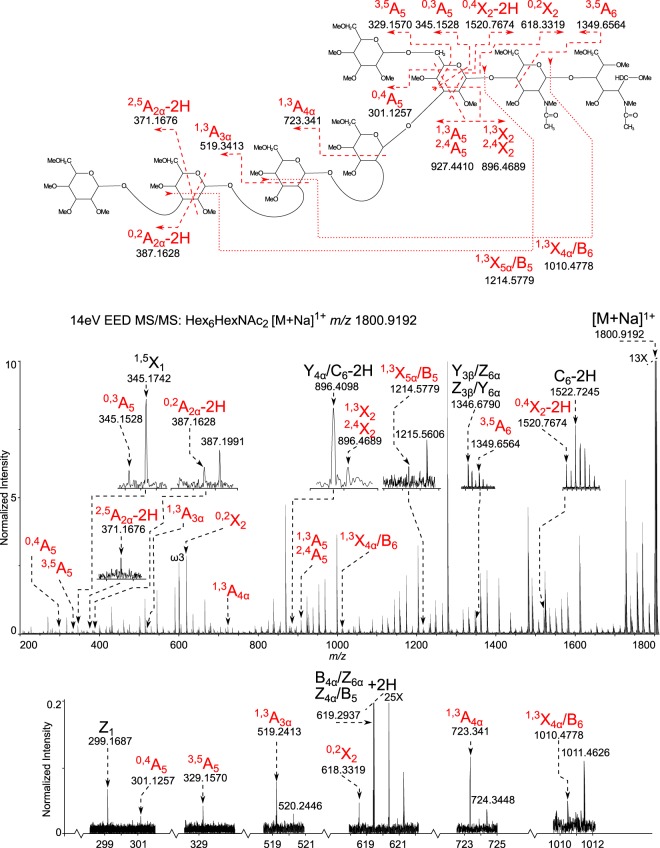
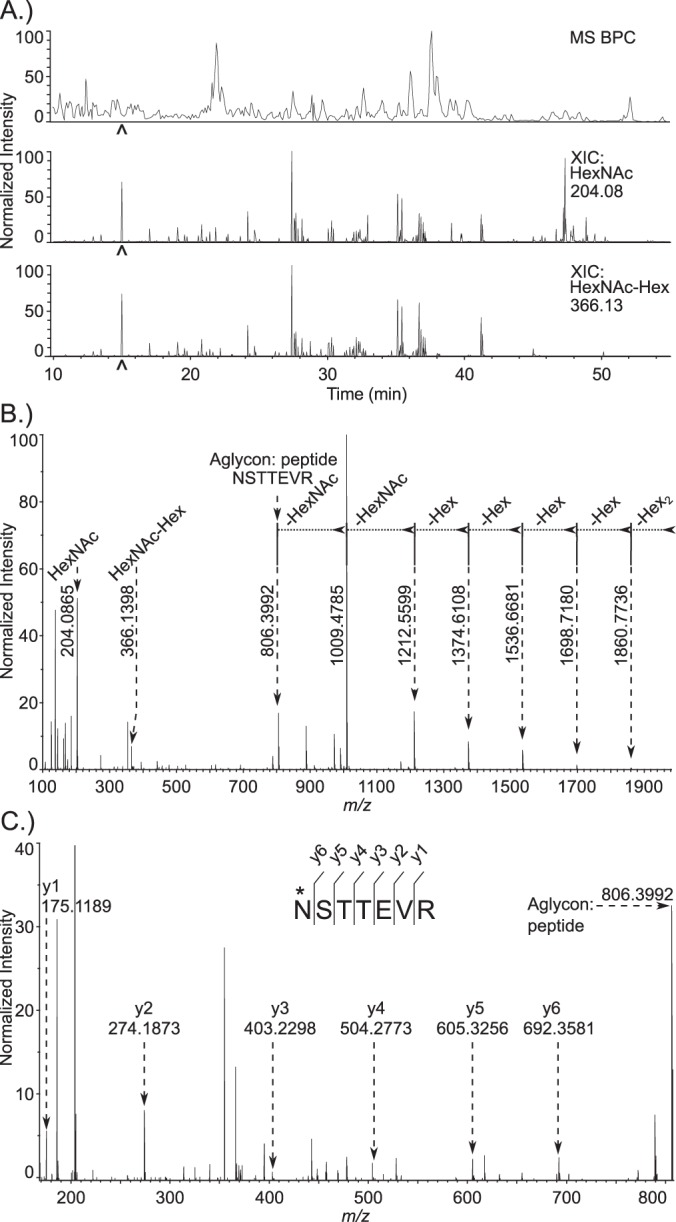
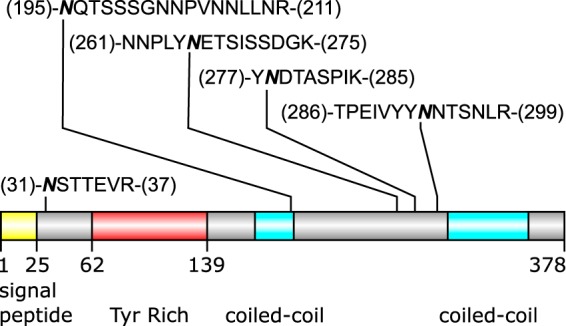
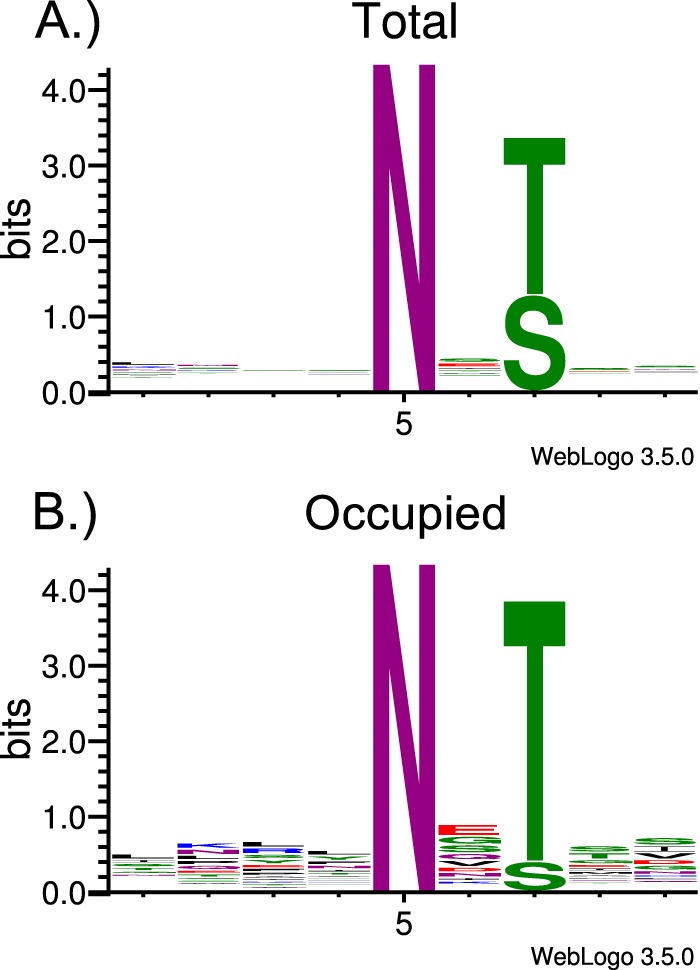
Cryptosporidium parvum causes severe diarrhea in infants in developing countries and in immunosuppressed persons, including those with AIDS. We are interested in the Asn-linked glycans (N-glycans) of C. parvum, because (1) the N-glycan precursor is predicted to contain five mannose and two glucose residues on a single long arm versus nine mannose and three glucose residues on the three-armed structure common in host N-glycans, (2) C. parvum is a rare eukaryote that lacks the machinery for N-glycan-dependent quality control of protein folding in the lumen of the Endoplasmic Reticulum (ER), and (3) ER and Golgi mannosidases, as well as glycosyltransferases that build complex N-glycans, are absent from the predicted proteome. The C. parvum N-glycans reported here, which were determined using a combination of collision-induced dissociation and electronic excitation dissociation, contain a single, unprocessed mannose arm ± terminal glucose on the trimannosyl chitobiose core. Upon nanoUPLC-MS/MS separation and analysis of the C. parvum tryptic peptides, the total ion and extracted oxonium ion chromatograms delineated 32 peptides with occupied N-glycan sites; these were derived from 16 glycoproteins. Although the number of potential N-glycan sites with Thr (NxT) is only about twice that with Ser (NxS), almost 90% of the occupied N-glycan sites contain NxT. The two most abundant C. parvum proteins modified with N-glycans were an immunodominant antigen on the surface of sporozoites (gp900) and the possible oocyst wall protein 1 (POWP1). Seven other glycoproteins with N-glycans were unique to C. parvum; five shared common ancestry with other apicomplexans; two glycoproteins shared common ancestry with many organisms. In summary, C. parvum N-glycans are remarkable for the absence of ER and Golgi modification and for the strong bias toward occupancy of N-glycan motifs containing Thr.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Checkley W., White A. C. Jr, Jaganath D., Arrowood M. J., Chalmers R. M., Chen X. M., Fayer R., Griffiths J. K., Guerrant R. L., Hedstrom L., Huston C. D., Kotloff K. L., Kang G., Mead J. R., Miller M., Petri W. A. Jr, Priest J. W., Roos D. S., Striepen B., Thompson R. C., Ward H. D., Van Voorhis W. A., Xiao L., Zhu G., and Houpt E. R. (2015) A review of the global burden, novel diagnostics, therapeutics, and vaccine targets for cryptosporidium. Lancet Infect. Dis. 15, 85–94 - PMC - PubMed
-
- Fayer R., and Xiao L. (2007) Cryptosporidium and cryptosporidiosis, CRC press
-
- Baldursson S., and Karanis P. (2011) Waterborne transmission of protozoan parasites: review of worldwide outbreaks - an update 2004–2010. Water Res. 45, 6603–6614 - PubMed
-
- Kotloff K. L., Nataro J. P., Blackwelder W. C., Nasrin D., Farag T. H., Panchalingam S., Wu Y., Sow S. O., Sur D., Breiman R. F., Faruque A. S., Zaidi A. K., Saha D., Alonso P. L., Tamboura B., Sanogo D., Onwuchekwa U., Manna B., Ramamurthy T., Kanungo S., Ochieng J. B., Omore R., Oundo J. O., Hossain A., Das S. K., Ahmed S., Qureshi S., Quadri F., Adegbola R. A., Antonio M., Hossain M. J., Akinsola A., Mandomando I., Nhampossa T., Acacio S., Biswas K., O'Reilly C. E., Mintz E. D., Berkeley L. Y., Muhsen K., Sommerfelt H., Robins-Browne R. M., and Levine M. M. (2013) Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): a prospective, case-control study. Lancet 382, 209–222 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
