

The evolution of the natural killer complex; a comparison between mammals using new high-quality genome assemblies and targeted annotation
- PMID: 28180967
- PMCID: PMC5350243
- DOI: 10.1007/s00251-017-0973-y
The evolution of the natural killer complex; a comparison between mammals using new high-quality genome assemblies and targeted annotation
Abstract
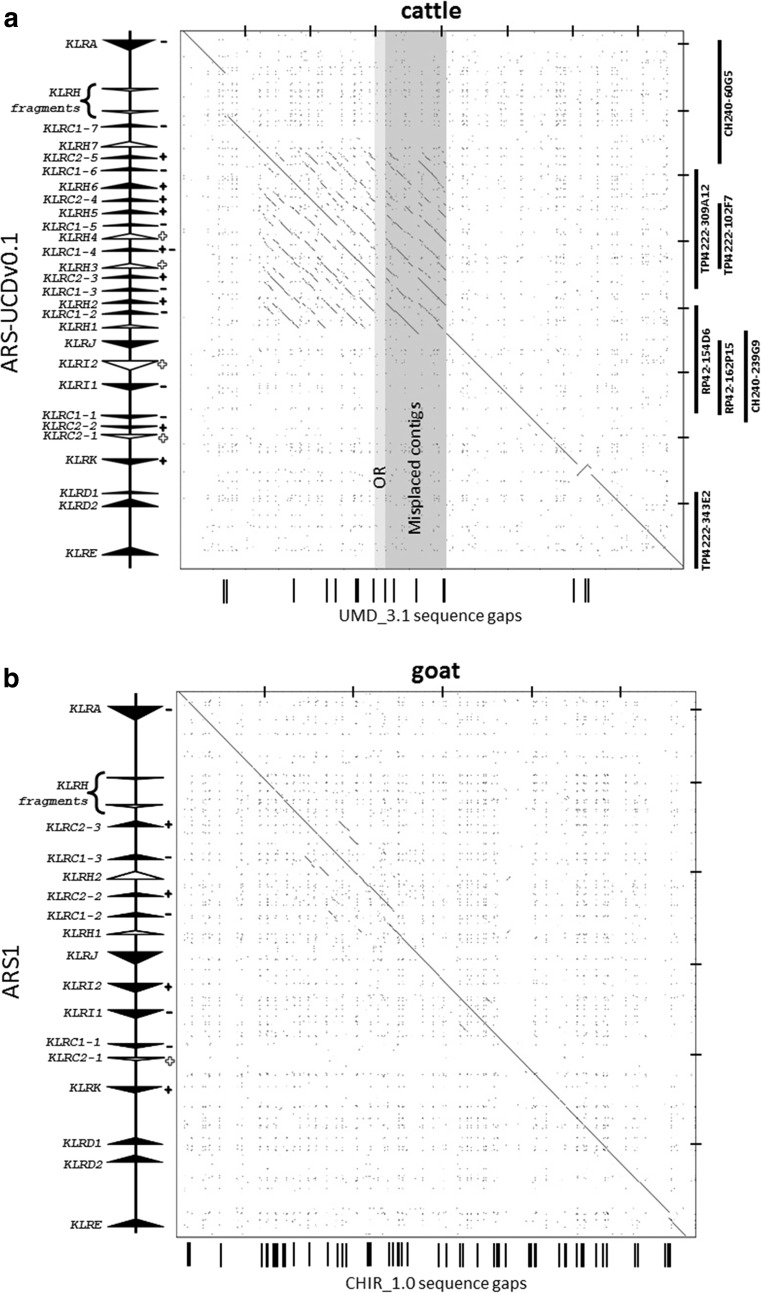
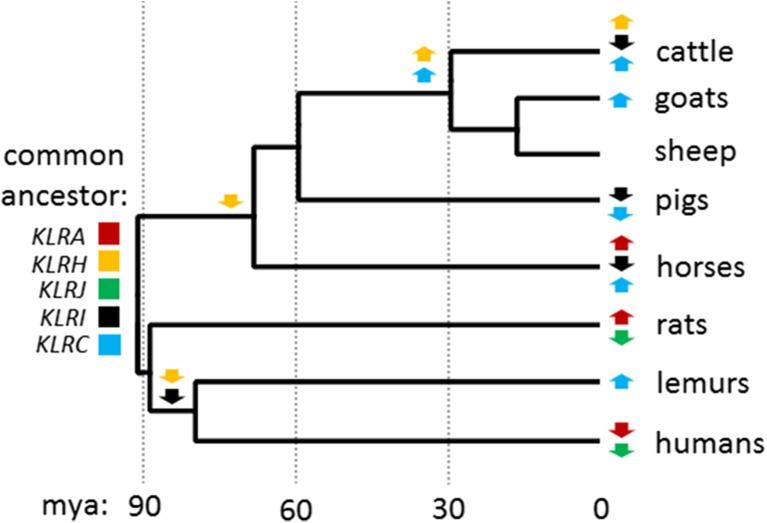
Natural killer (NK) cells are a diverse population of lymphocytes with a range of biological roles including essential immune functions. NK cell diversity is in part created by the differential expression of cell surface receptors which modulate activation and function, including multiple subfamilies of C-type lectin receptors encoded within the NK complex (NKC). Little is known about the gene content of the NKC beyond rodent and primate lineages, other than it appears to be extremely variable between mammalian groups. We compared the NKC structure between mammalian species using new high-quality draft genome assemblies for cattle and goat; re-annotated sheep, pig, and horse genome assemblies; and the published human, rat, and mouse lemur NKC. The major NKC genes are largely in the equivalent positions in all eight species, with significant independent expansions and deletions between species, allowing us to propose a model for NKC evolution during mammalian radiation. The ruminant species, cattle and goats, have independently evolved a second KLRC locus flanked by KLRA and KLRJ, and a novel KLRH-like gene has acquired an activating tail. This novel gene has duplicated several times within cattle, while other activating receptor genes have been selectively disrupted. Targeted genome enrichment in cattle identified varying levels of allelic polymorphism between the NKC genes concentrated in the predicted extracellular ligand-binding domains. This novel recombination and allelic polymorphism is consistent with NKC evolution under balancing selection, suggesting that this diversity influences individual immune responses and may impact on differential outcomes of pathogen infection and vaccination.
Keywords: C-type lectin; KLRA; KLRC; Leukocyte receptor complex; Natural killer cells; Natural killer complex.
Figures



References
-
- Averdam A, Petersen B, Rosner C, Neff J, Roos C, Eberle M, Aujard F, Münch C, Schempp W, Carrington M, Shiina T, Inoko H, Knaust F, Coggill P, Sehra H, Beck S, Abi-Rached L, Reinhardt R, Walter L. A novel system of polymorphic and diverse NK cell receptors in primates. PLoS Genet. 2009;5:e1000688. doi: 10.1371/journal.pgen.1000688. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
