

Coordinate activities of BRD4 and CDK9 in the transcriptional elongation complex are required for TGFβ-induced Nox4 expression and myofibroblast transdifferentiation
- PMID: 28182006
- PMCID: PMC5386453
- DOI: 10.1038/cddis.2016.434
Coordinate activities of BRD4 and CDK9 in the transcriptional elongation complex are required for TGFβ-induced Nox4 expression and myofibroblast transdifferentiation
Abstract
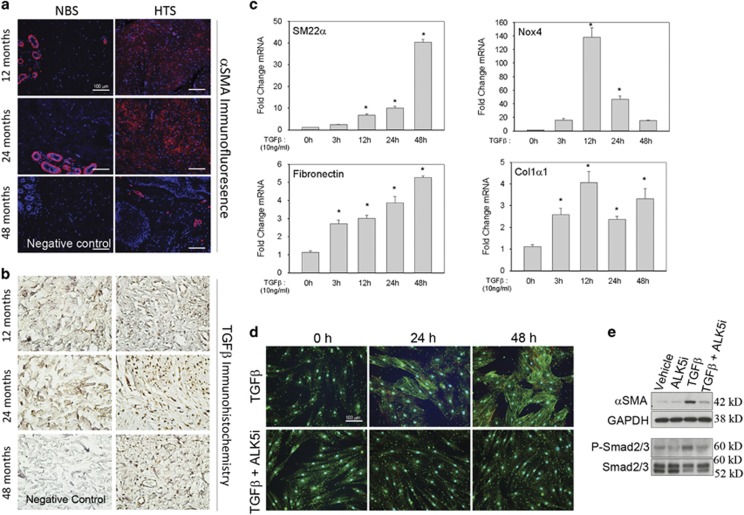
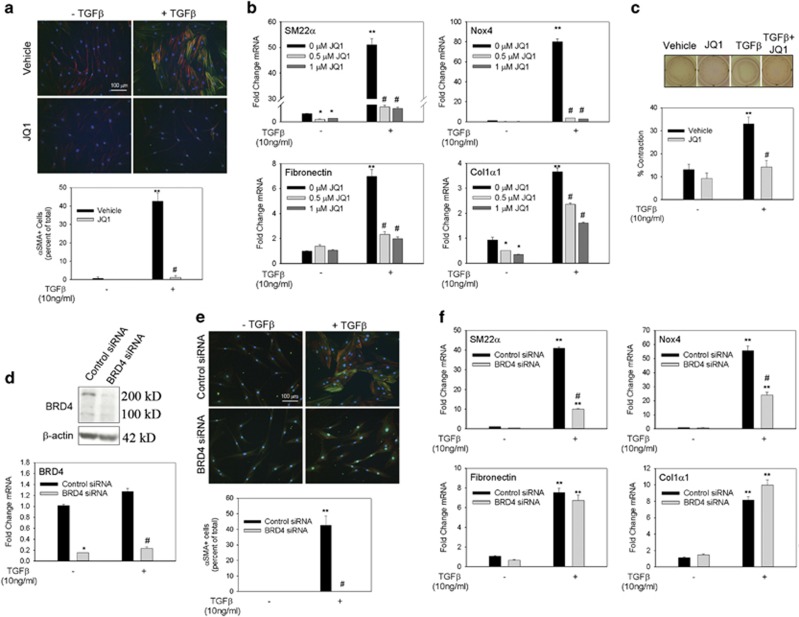
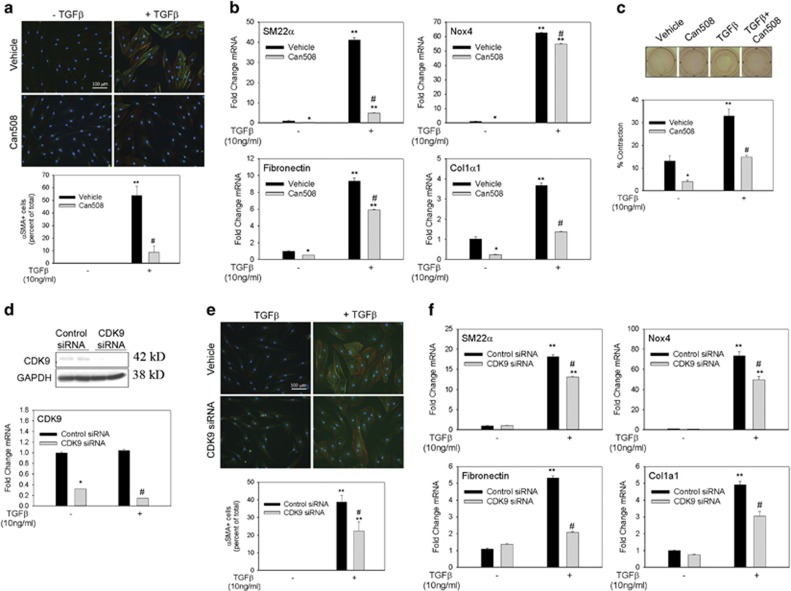
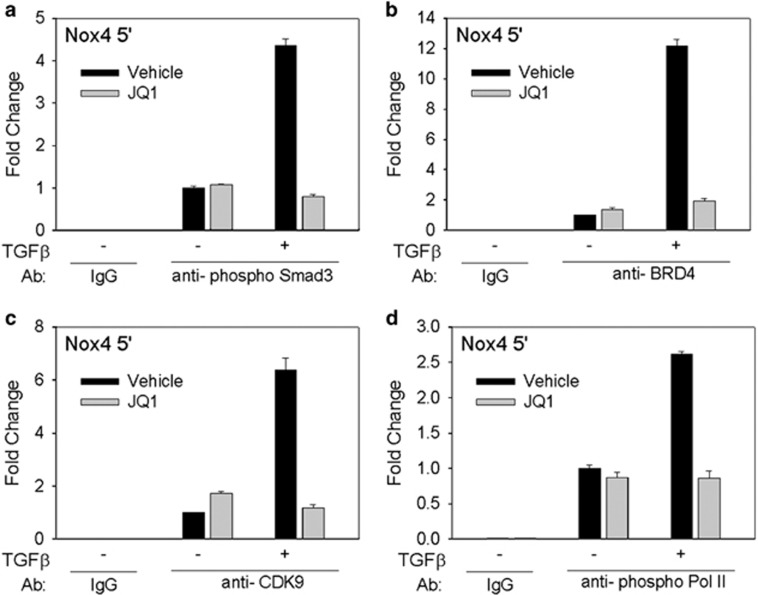
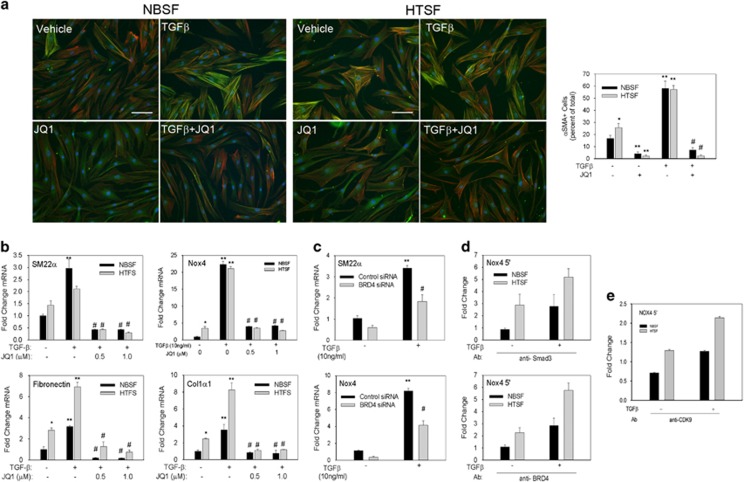
Transdifferentiation of quiescent dermal fibroblasts to secretory myofibroblasts has a central role in wound healing and pathological scar formation. This myofibroblast transdifferentiation process involves TGFβ-induced de novo synthesis of alpha smooth muscle cell actin (αSMA)+ fibers that enhance contractility as well as increased expression of extracellular matrix (ECM) proteins, including collagen and fibronectin. These processes are mediated upstream by the reactive oxygen species (ROS)-producing enzyme Nox4, whose induction by TGFβ is incompletely understood. In this study, we demonstrate that Nox4 is involved in αSMA+ fiber formation and collagen production in primary human dermal fibroblasts (hDFs) using a small-molecule inhibitor and siRNA-mediated silencing. Furthermore, TGFβ-induced signaling via Smad3 is required for myofibroblast transformation and Nox4 upregulation. Immunoprecipitation-selected reaction monitoring (IP-SRM) assays of the activated Smad3 complex suggest that it couples with the epigenetic reader and transcription co-activator bromodomain and extraterminal (BET) domain containing protein 4 (BRD4) to promote Nox4 transcription. In addition, cyclin-dependent kinase 9 (CDK9), a component of positive transcription elongation factor, binds to BRD4 after TGFβ stimulation and is also required for RNA polymerase II phosphorylation and Nox4 transcription regulation. Surprisingly, BRD4 depletion decreases myofibroblast differentiation but does not affect collagen or fibronectin expression in primary skin fibroblasts, whereas knockdown of CDK9 decreases all myofibroblast genes. We observe enhanced numbers and persistence of myofibroblast formation and TGFβ signaling in hypertrophic scars. BRD4 inhibition reverses hypertrophic skin fibroblast transdifferentiation to myofibroblasts. Our data indicate that BRD4 and CDK9 have independent, coordinated roles in promoting the myofibroblast transition and suggest that inhibition of the Smad3-BRD4 pathway may be a useful strategy to limit hypertrophic scar formation after burn injury.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Similar articles
-
Ligand-activated PPARδ upregulates α-smooth muscle actin expression in human dermal fibroblasts: A potential role for PPARδ in wound healing.J Dermatol Sci. 2015 Dec;80(3):186-95. doi: 10.1016/j.jdermsci.2015.10.005. Epub 2015 Oct 9. J Dermatol Sci. 2015. PMID: 26481780
-
Targeting of NADPH oxidase in vitro and in vivo suppresses fibroblast activation and experimental skin fibrosis.Exp Dermatol. 2017 Jan;26(1):73-81. doi: 10.1111/exd.13180. Exp Dermatol. 2017. PMID: 27576129
-
NAD(P)H oxidase mediates TGF-beta1-induced activation of kidney myofibroblasts.J Am Soc Nephrol. 2010 Jan;21(1):93-102. doi: 10.1681/ASN.2009020146. Epub 2009 Nov 19. J Am Soc Nephrol. 2010. PMID: 19926889 Free PMC article.
-
Myofibroblast transdifferentiation: The dark force in ocular wound healing and fibrosis.Prog Retin Eye Res. 2017 Sep;60:44-65. doi: 10.1016/j.preteyeres.2017.08.001. Epub 2017 Aug 12. Prog Retin Eye Res. 2017. PMID: 28807717 Free PMC article. Review.
-
RSV Reprograms the CDK9•BRD4 Chromatin Remodeling Complex to Couple Innate Inflammation to Airway Remodeling.Viruses. 2020 Apr 22;12(4):472. doi: 10.3390/v12040472. Viruses. 2020. PMID: 32331282 Free PMC article. Review.
Cited by
-
Discovery of RSV-Induced BRD4 Protein Interactions Using Native Immunoprecipitation and Parallel Accumulation-Serial Fragmentation (PASEF) Mass Spectrometry.Viruses. 2021 Mar 11;13(3):454. doi: 10.3390/v13030454. Viruses. 2021. PMID: 33799525 Free PMC article.
-
Delivery of a BET protein degrader via a CEACAM6-targeted antibody-drug conjugate inhibits tumour growth in pancreatic cancer models.Nat Commun. 2024 Mar 11;15(1):2192. doi: 10.1038/s41467-024-46167-1. Nat Commun. 2024. PMID: 38467634 Free PMC article.
-
BRD4 inhibition for the treatment of pathological organ fibrosis.F1000Res. 2017 Jun 28;6:F1000 Faculty Rev-1015. doi: 10.12688/f1000research.11339.1. eCollection 2017. F1000Res. 2017. PMID: 28721198 Free PMC article. Review.
-
CXCL6-EGFR-induced Kupffer cells secrete TGF-β1 promoting hepatic stellate cell activation via the SMAD2/BRD4/C-MYC/EZH2 pathway in liver fibrosis.J Cell Mol Med. 2018 Oct;22(10):5050-5061. doi: 10.1111/jcmm.13787. Epub 2018 Aug 14. J Cell Mol Med. 2018. PMID: 30106235 Free PMC article.
-
Epigenetics in Cardiac Fibrosis: Emphasis on Inflammation and Fibroblast Activation.JACC Basic Transl Sci. 2018 Nov 12;3(5):704-715. doi: 10.1016/j.jacbts.2018.05.003. eCollection 2018 Oct. JACC Basic Transl Sci. 2018. PMID: 30456341 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
