

Deciphering the molecular mechanism responsible for GCaMP6m's Ca2+-dependent change in fluorescence
- PMID: 28182677
- PMCID: PMC5300113
- DOI: 10.1371/journal.pone.0170934
Deciphering the molecular mechanism responsible for GCaMP6m's Ca2+-dependent change in fluorescence
Abstract
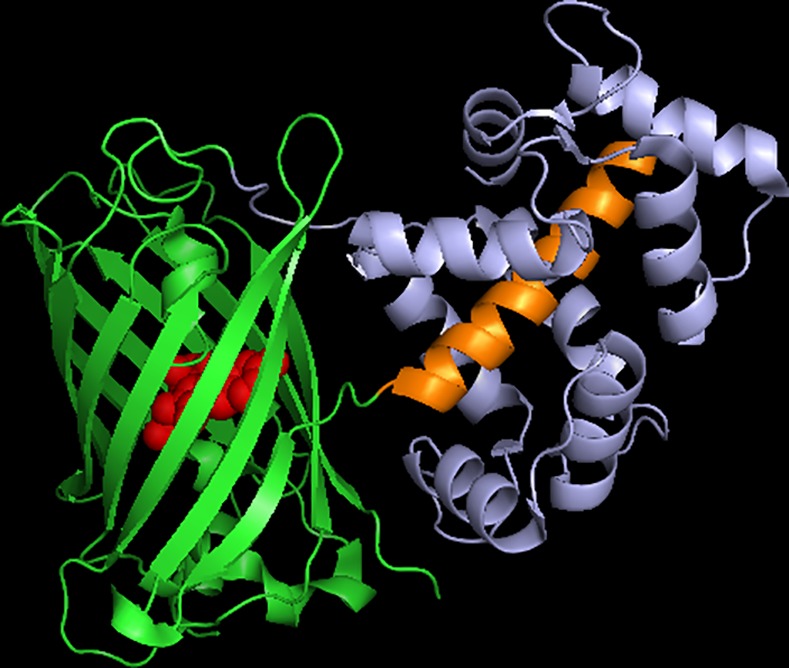
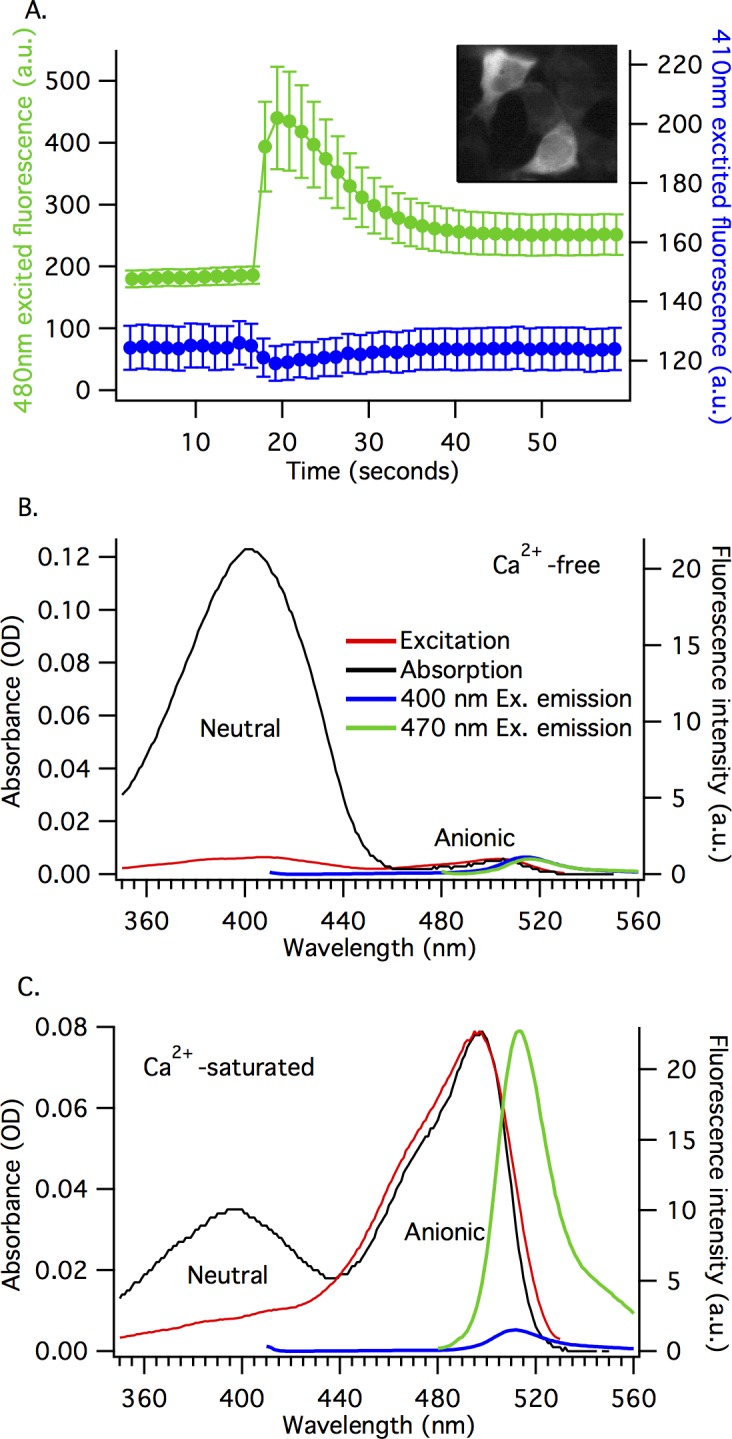
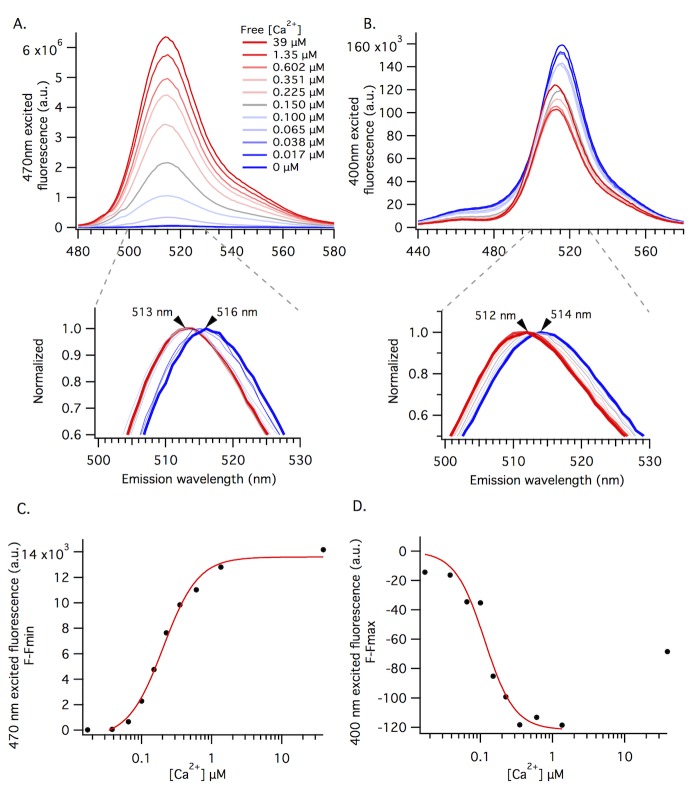
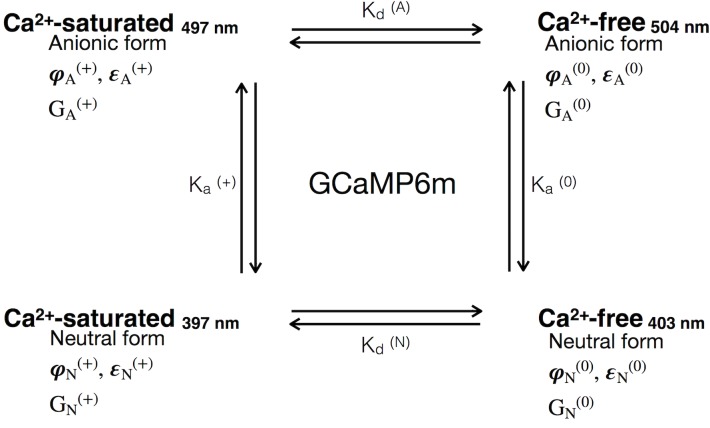
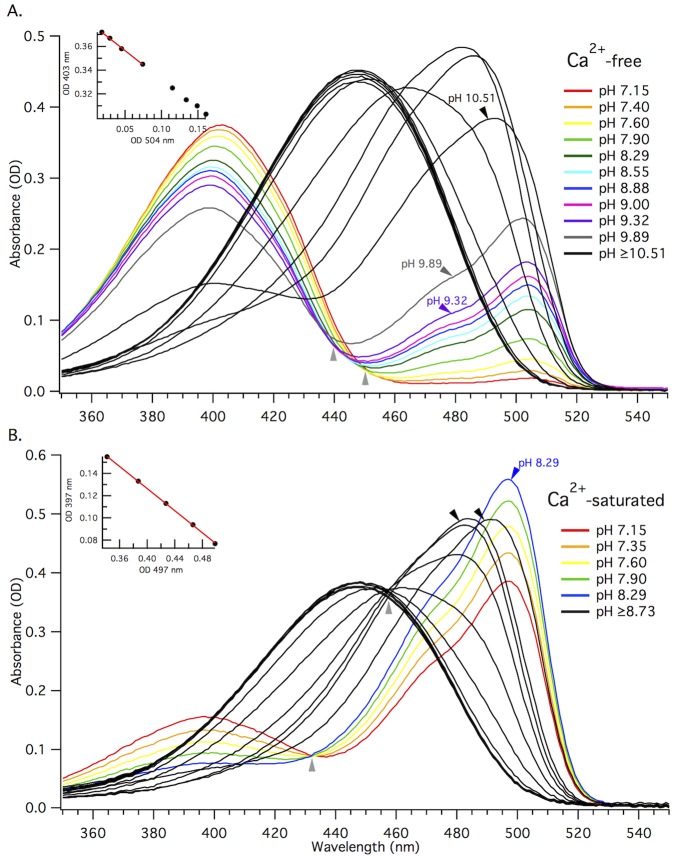
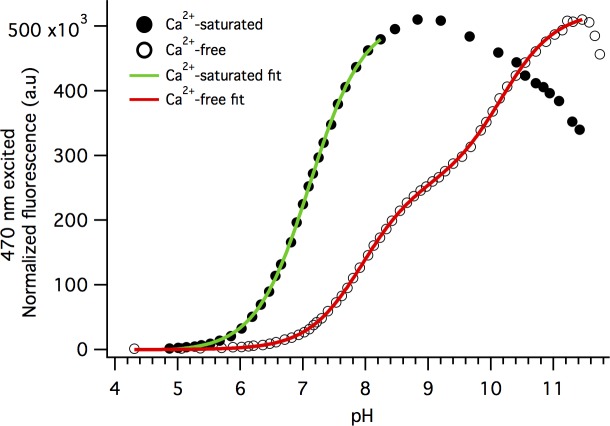
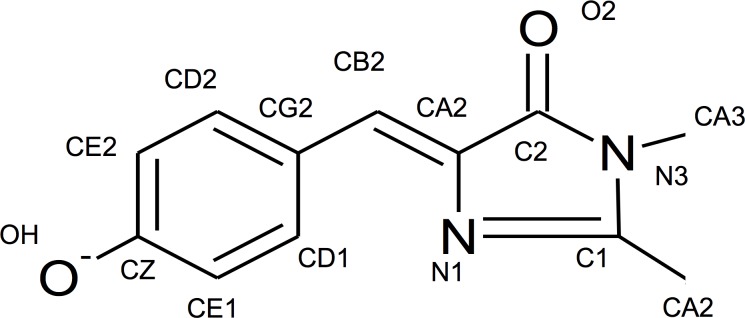
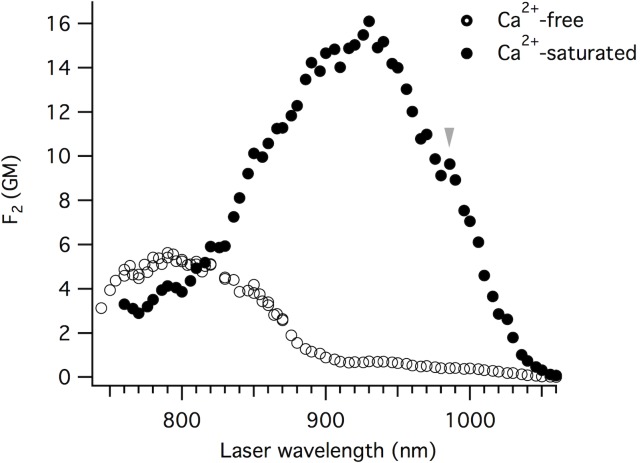
The goal of this work is to determine how GCaMP6m's fluorescence is altered in response to Ca2+-binding. Our detailed spectroscopic study reveals the simplest explanation for how GCaMP6m changes fluorescence in response to Ca2+ is with a four-state model, in which a Ca2+-dependent change of the chromophore protonation state, due to a shift in pKa, is the predominant factor. The pKa shift is quantitatively explained by a change in electrostatic potential around the chromophore due to the conformational changes that occur in the protein when calmodulin binds Ca2+ and interacts with the M13 peptide. The absolute pKa values for the Ca2+-free and Ca2+-saturated states of GCaMP6m are critical to its high signal-to-noise ratio. This mechanism has important implications for further improvements to GCaMP6m and potentially for other similarly designed biosensors.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
