

Zika Virus Pathogenesis and Tissue Tropism
- PMID: 28182948
- PMCID: PMC5328190
- DOI: 10.1016/j.chom.2017.01.004
Zika Virus Pathogenesis and Tissue Tropism
Abstract
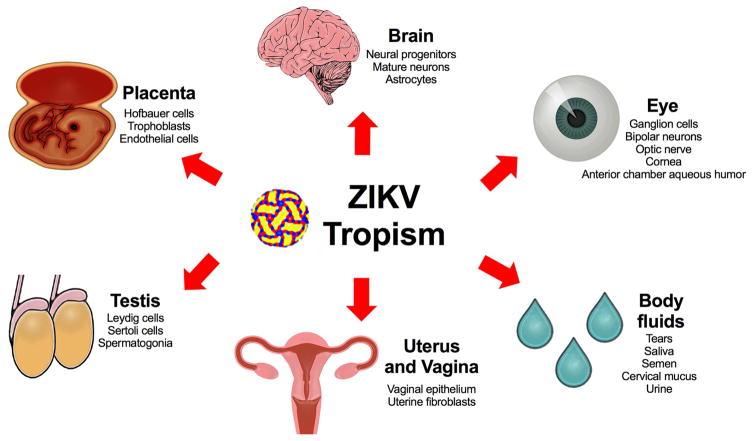
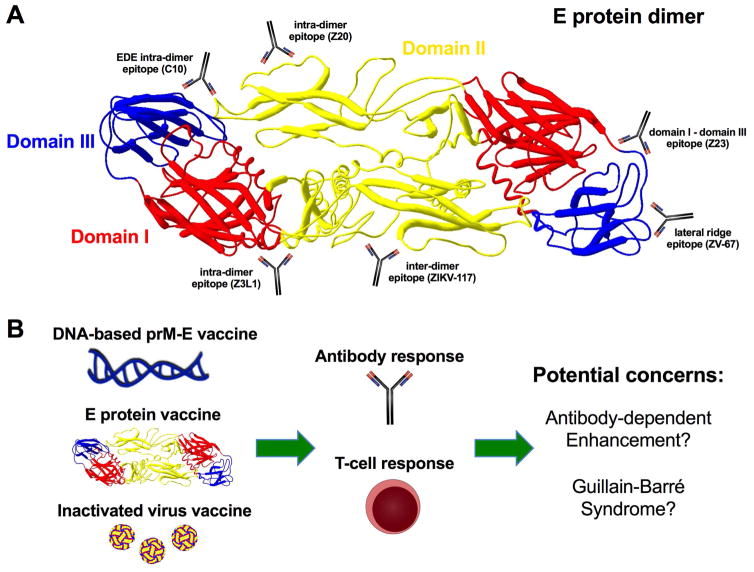
Although Zika virus (ZIKV) was isolated approximately 70 years ago, few experimental studies had been published prior to 2016. The recent spread of ZIKV to countries in the Western Hemisphere is associated with reports of microcephaly, congenital malformations, and Guillain-Barré syndrome. This has resulted in ZIKV being declared a public health emergency and has greatly accelerated the pace of ZIKV research and discovery. Within a short time period, useful mouse and non-human primate disease models have been established, and pre-clinical evaluation of therapeutics and vaccines has begun. Unexpectedly, ZIKV exhibits a broad tropism and persistence in body tissues and fluids, which contributes to the clinical manifestations and epidemiology that have been observed during the current epidemic. In this Review, we highlight recent advances in our understanding of ZIKV pathogenesis, tissue tropism, and the resulting pathology and discuss areas for future investigation.
Keywords: Zika virus; congenital infection; flavivirus; host immunity; innate immunity; microcephaly; pathogenesis; tropism; viral persistence.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
