

CLUH regulates mitochondrial metabolism by controlling translation and decay of target mRNAs
- PMID: 28188211
- PMCID: PMC5350512
- DOI: 10.1083/jcb.201607019
CLUH regulates mitochondrial metabolism by controlling translation and decay of target mRNAs
Abstract
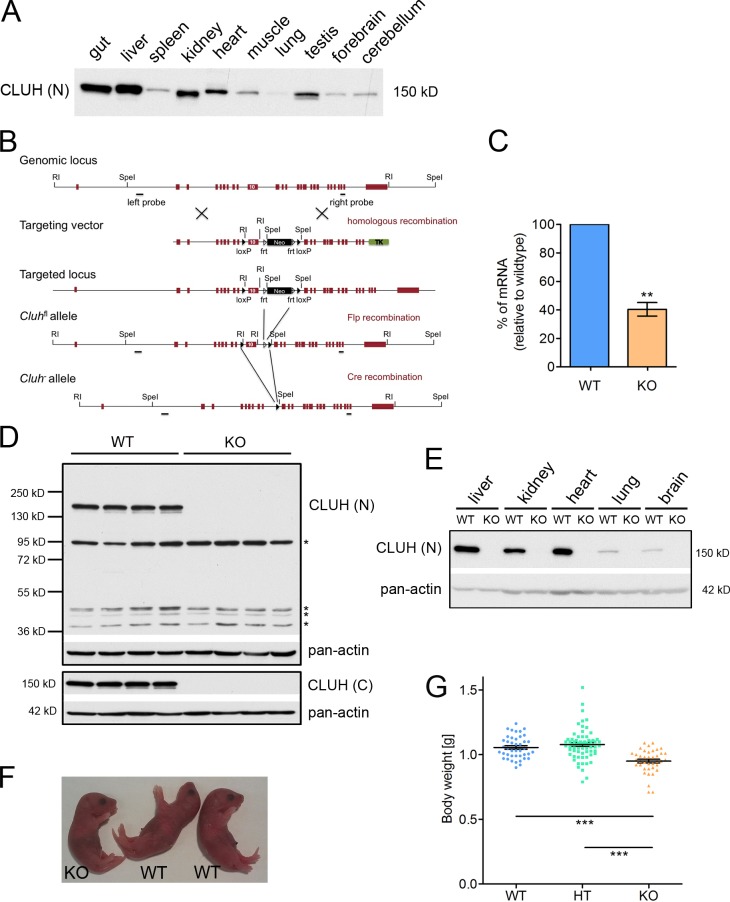
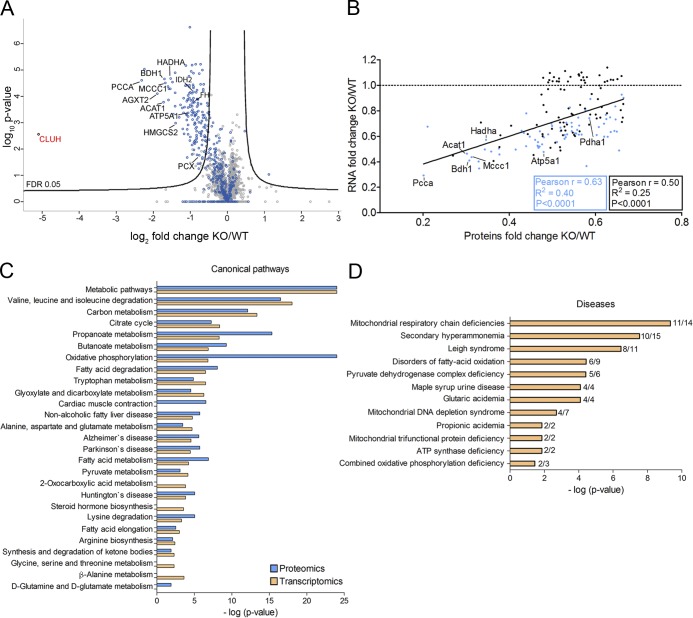
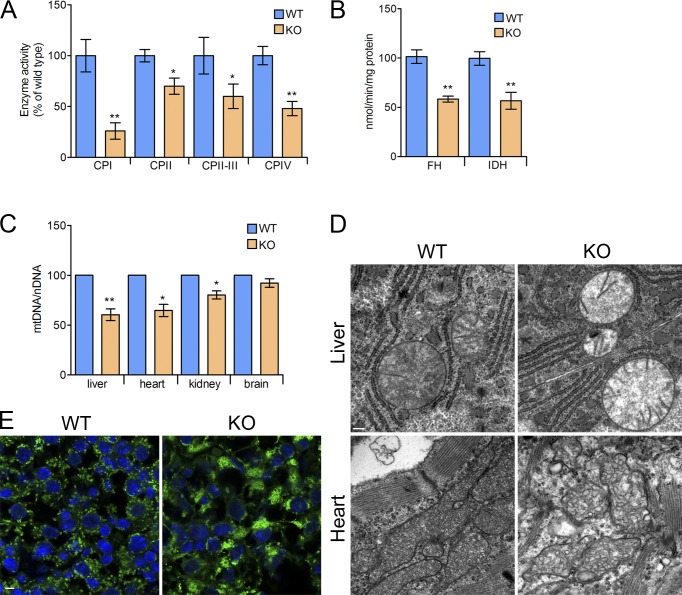
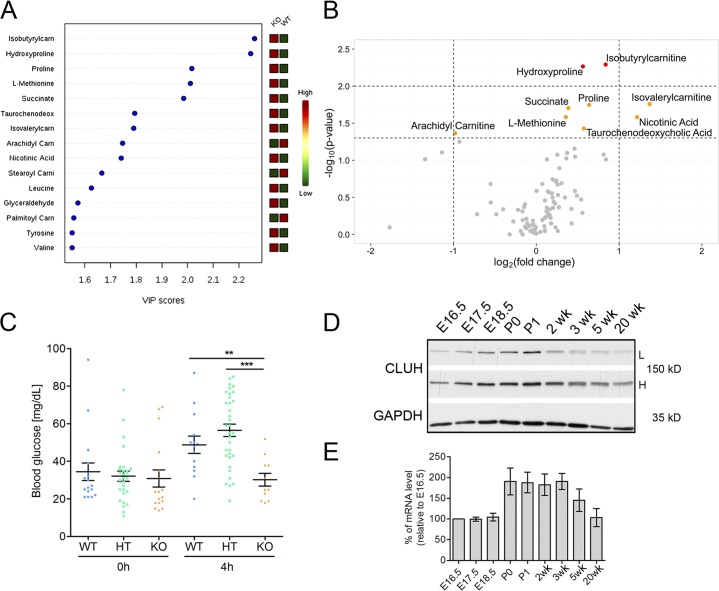
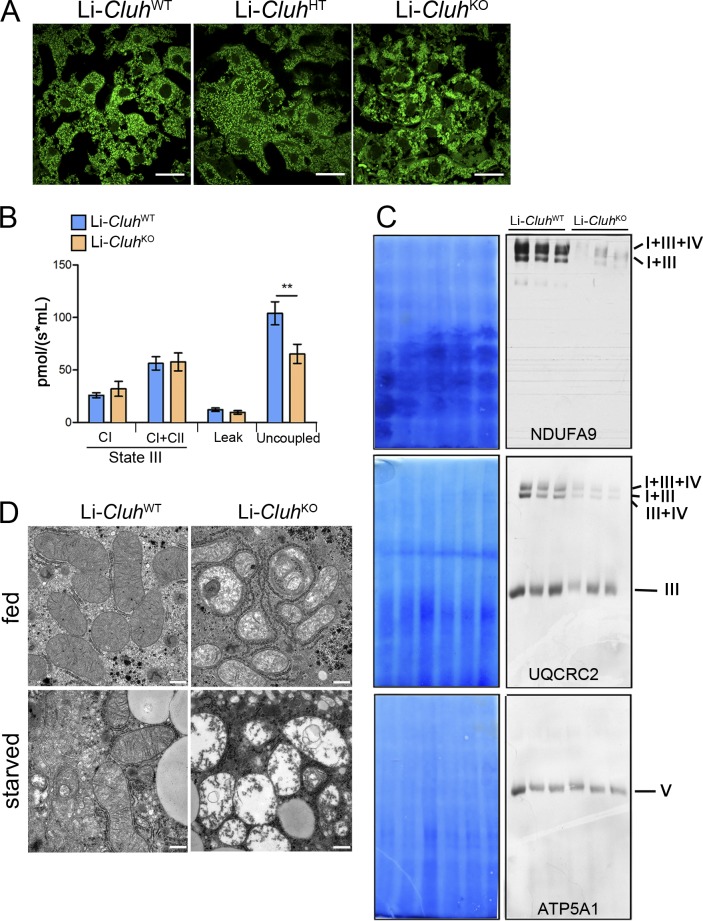
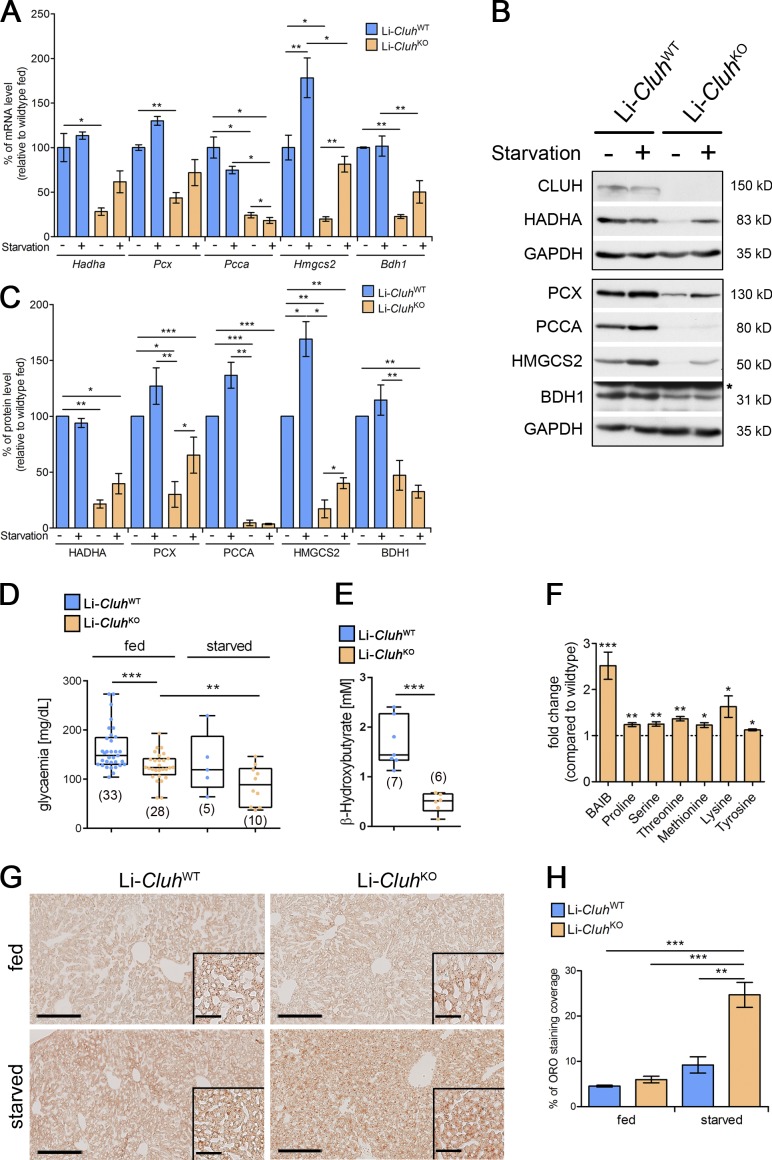
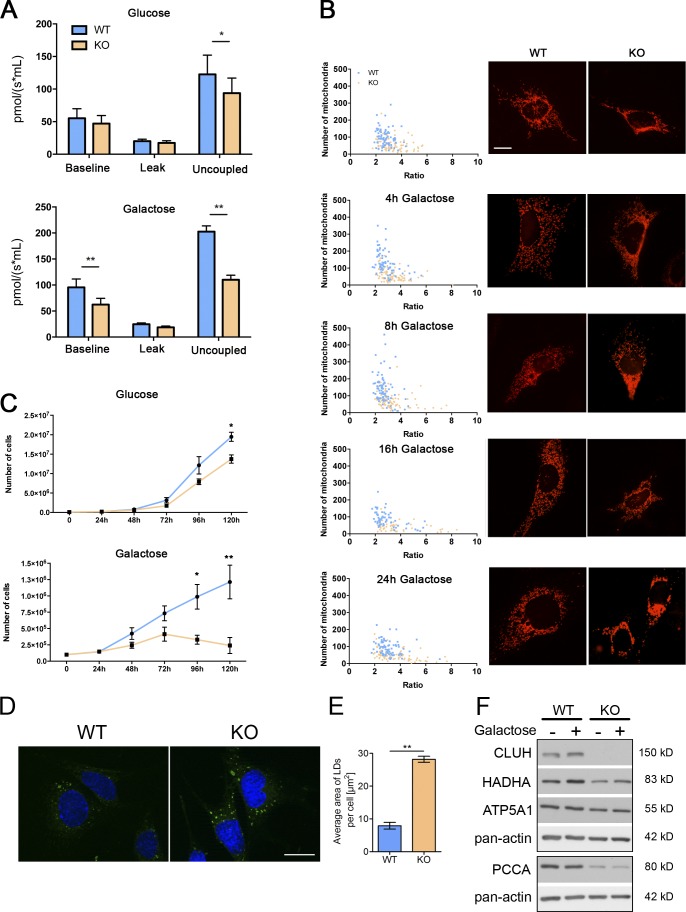
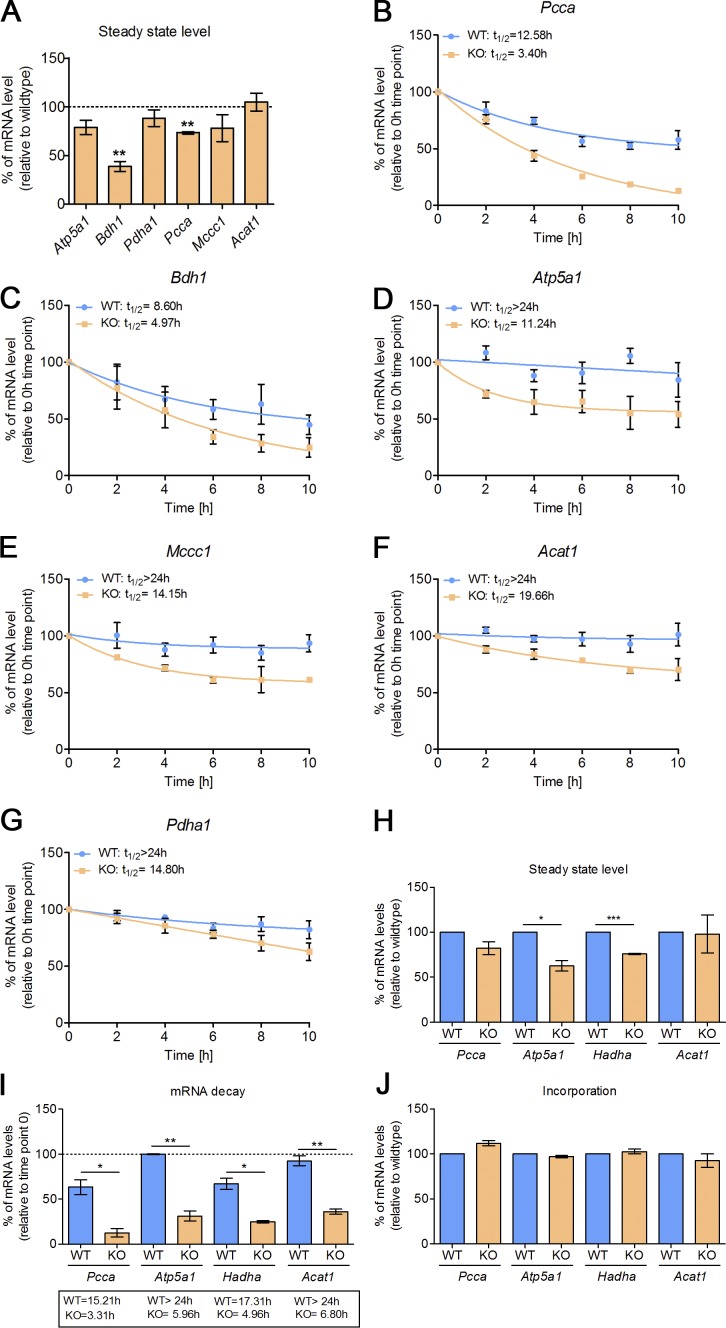
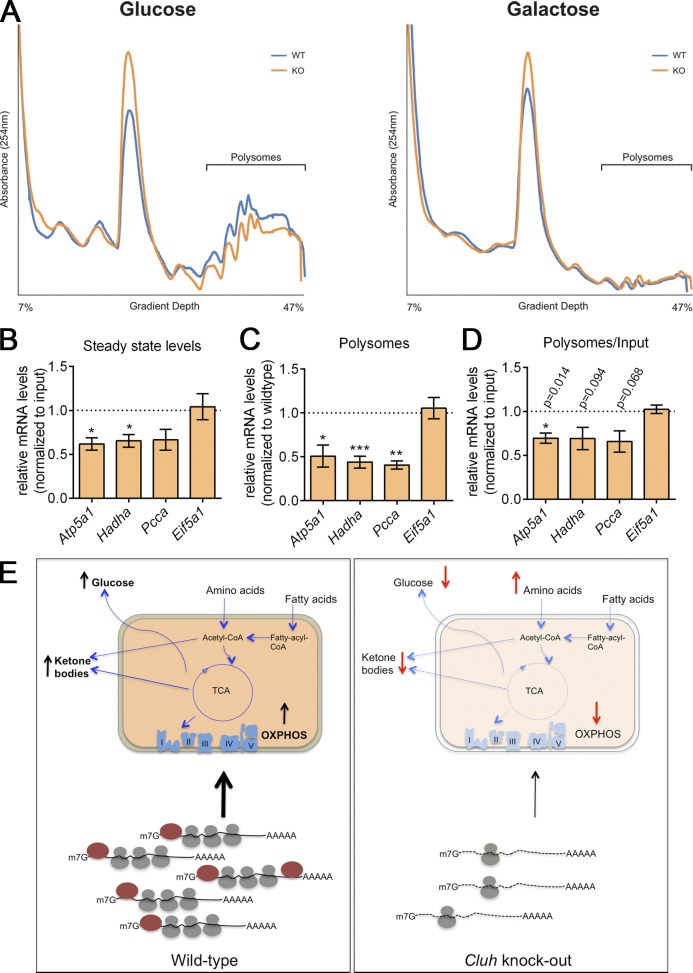
Mitochondria are essential organelles that host crucial metabolic pathways and produce adenosine triphosphate. The mitochondrial proteome is heterogeneous among tissues and can dynamically change in response to different metabolic conditions. Although the transcriptional programs that govern mitochondrial biogenesis and respiratory function are well known, posttranscriptional regulatory mechanisms remain unclear. In this study, we show that the cytosolic RNA-binding protein clustered mitochondria homologue (CLUH) regulates the expression of a mitochondrial protein network supporting key metabolic programs required under nutrient deprivation. CLUH exerts its function by controlling the stability and translation of target messenger RNAs. In the absence of Cluh, mitochondria are severely depleted of crucial enzymes involved in catabolic energy-converting pathways. CLUH preserves oxidative mitochondrial function and glucose homeostasis, thus preventing death at the fetal-neonatal transition. In the adult liver, CLUH ensures maximal respiration capacity and the metabolic response to starvation. Our results shed new light on the posttranscriptional mechanisms controlling the expression of mitochondrial proteins and suggest novel strategies to tailor mitochondrial function to physiological and pathological conditions.
© 2017 Schatton et al.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
