

Amygdalar Gating of Early Sensory Processing through Interactions with Locus Coeruleus
- PMID: 28188216
- PMCID: PMC5354340
- DOI: 10.1523/JNEUROSCI.2797-16.2017
Amygdalar Gating of Early Sensory Processing through Interactions with Locus Coeruleus
Abstract
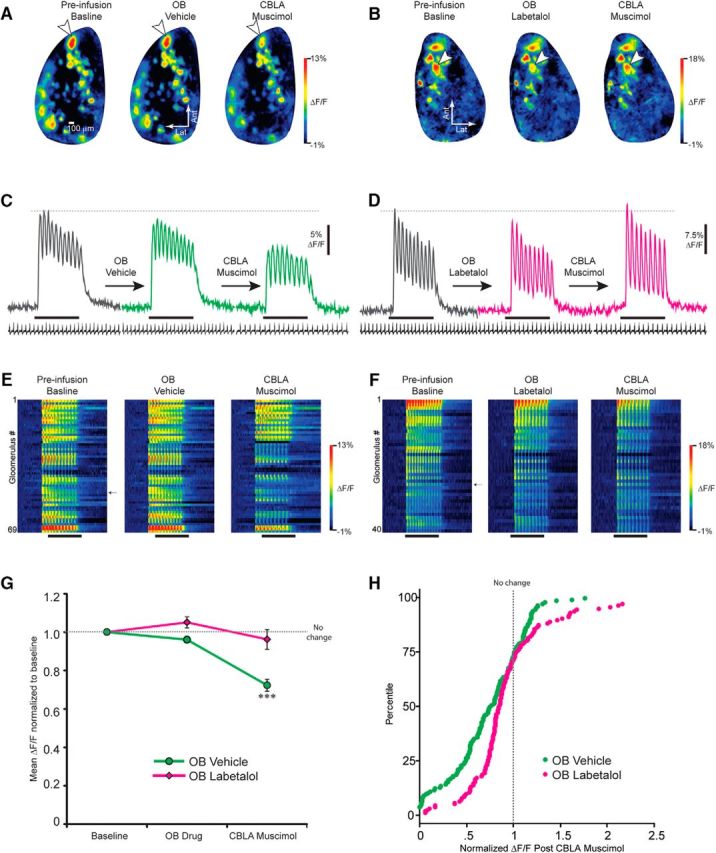
Fear- and stress-induced activity in the amygdala has been hypothesized to influence sensory brain regions through the influence of the amygdala on neuromodulatory centers. To directly examine this relationship, we used optical imaging to observe odor-evoked activity in populations of olfactory bulb inhibitory interneurons and of synaptic terminals of olfactory sensory neurons (the primary sensory neurons of the olfactory system, which provide the initial olfactory input to the brain) during pharmacological inactivation of amygdala and locus coeruleus (LC) in mice. Although the amygdala does not directly project to the olfactory bulb, joint pharmacological inactivation of the central, basolateral, and lateral nuclei of the amygdala nonetheless strongly suppressed odor-evoked activity in GABAergic inhibitory interneuron populations in the OB. This suppression was prevented by inactivation of LC or pretreatment of the olfactory bulb with a broad-spectrum noradrenergic receptor antagonist. Visualization of synaptic output from olfactory sensory neuron terminals into the olfactory bulb of the brain revealed that amygdalar inactivation preferentially strengthened the odor-evoked synaptic output of weakly activated populations of sensory afferents from the nose, thus demonstrating a change in sensory gating potentially mediated by local inhibition of olfactory sensory neuron terminals. We conclude that amygdalar activity influences olfactory processing as early as the primary sensory input to the brain by modulating norepinephrine release from the locus coeruleus into the olfactory bulb. These findings show that the amygdala and LC state actively determines which sensory signals are selected for processing in sensory brain regions. Similar local circuitry operates in the olfactory, visual, and auditory systems, suggesting a potentially shared mechanism across modalities.SIGNIFICANCE STATEMENT The affective state is increasingly understood to influence early neural processing of sensory stimuli, not just the behavioral response to those stimuli. The present study elucidates one circuit by which the amygdala, a critical structure for emotional learning, valence coding, and stress, can shape sensory input to the brain and early sensory processing through its connections to the locus coeruleus. One function of this interaction appears to be sensory gating, because inactivating the central, basolateral, and lateral nuclei of the amygdala selectively strengthened the weakest olfactory inputs to the brain. This linkage of amygdalar and LC output to primary sensory signaling may have implications for affective disorders that include sensory dysfunctions like hypervigilance, attentional bias, and impaired sensory gating.
Keywords: affective; amygdala; emotional; locus coeruleus; olfactory; sensory gating.
Copyright © 2017 the authors 0270-6474/17/373085-17$15.00/0.
Figures







Similar articles
-
Noradrenergic plasticity of olfactory sensory neuron inputs to the main olfactory bulb.J Neurosci. 2014 Nov 12;34(46):15234-43. doi: 10.1523/JNEUROSCI.0551-14.2014. J Neurosci. 2014. PMID: 25392492 Free PMC article.
-
Early-life stress disrupts attachment learning: the role of amygdala corticosterone, locus ceruleus corticotropin releasing hormone, and olfactory bulb norepinephrine.J Neurosci. 2009 Dec 16;29(50):15745-55. doi: 10.1523/JNEUROSCI.4106-09.2009. J Neurosci. 2009. PMID: 20016090 Free PMC article.
-
Locus coeruleus stimulation modulates olfactory bulb evoked potentials.Brain Res Bull. 1987 Jun;18(6):767-70. doi: 10.1016/0361-9230(87)90213-9. Brain Res Bull. 1987. PMID: 3621000
-
Construction of odor representations by olfactory bulb microcircuits.Prog Brain Res. 2014;208:177-203. doi: 10.1016/B978-0-444-63350-7.00007-3. Prog Brain Res. 2014. PMID: 24767483 Review.
-
Circuit formation and maintenance--perspectives from the mammalian olfactory bulb.Curr Opin Neurobiol. 2010 Feb;20(1):134-40. doi: 10.1016/j.conb.2009.11.001. Epub 2009 Dec 11. Curr Opin Neurobiol. 2010. PMID: 20005696 Review.
Cited by
-
Handling stress impairs learning through a mechanism involving caspase-1 activation and adenosine signaling.Brain Behav Immun. 2019 Aug;80:763-776. doi: 10.1016/j.bbi.2019.05.025. Epub 2019 May 17. Brain Behav Immun. 2019. PMID: 31108171 Free PMC article.
-
Age-Related Intrinsic Functional Connectivity Changes of Locus Coeruleus from Childhood to Older Adults.Brain Sci. 2021 Nov 10;11(11):1485. doi: 10.3390/brainsci11111485. Brain Sci. 2021. PMID: 34827484 Free PMC article.
-
NIH Workshop Report: sensory nutrition and disease.Am J Clin Nutr. 2021 Jan 4;113(1):232-245. doi: 10.1093/ajcn/nqaa302. Am J Clin Nutr. 2021. PMID: 33300030 Free PMC article.
-
A Systems Neuroscience Approach to Migraine.Neuron. 2018 Mar 7;97(5):1004-1021. doi: 10.1016/j.neuron.2018.01.029. Neuron. 2018. PMID: 29518355 Free PMC article. Review.
-
Respiratory entrainment of the locus coeruleus modulates arousal level to avoid physical risks from external vibration.Sci Rep. 2023 May 1;13(1):7069. doi: 10.1038/s41598-023-32995-6. Sci Rep. 2023. PMID: 37127727 Free PMC article.
References
-
- Akerboom J, Chen TW, Wardill TJ, Tian L, Marvin JS, Mutlu S, Calderón NC, Esposti F, Borghuis BG, Sun XR, Gordus A, Orger MB, Portugues R, Engert F, Macklin JJ, Filosa A, Aggarwal A, Kerr RA, Takagi R, Kracun S, et al. (2012) Optimization of a GCaMP calcium indicator for neural activity imaging. J Neurosci 32:13819–13840. 10.1523/JNEUROSCI.2601-12.2012 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous