

Microexons: discovery, regulation, and function
- PMID: 28188674
- PMCID: PMC5863539
- DOI: 10.1002/wrna.1418
Microexons: discovery, regulation, and function
Abstract
The importance of RNA splicing in numerous cellular processes is well established. However, an underappreciated aspect is the ability of the spliceosome to recognize a set of very small (3-30 nucleotide, 1-10 amino acid) exons named microexons. Despite their small size, microexons and their regulation through alternative splicing have now been shown to play critical roles in protein and system function. Here we review the discovery of microexons over time and the mechanisms by which their splicing is regulated, including recent progress made through deep RNA sequencing. We also discuss the functional role of microexons in biology and disease. WIREs RNA 2017, 8:e1418. doi: 10.1002/wrna.1418 For further resources related to this article, please visit the WIREs website.
© 2017 Wiley Periodicals, Inc.
Figures
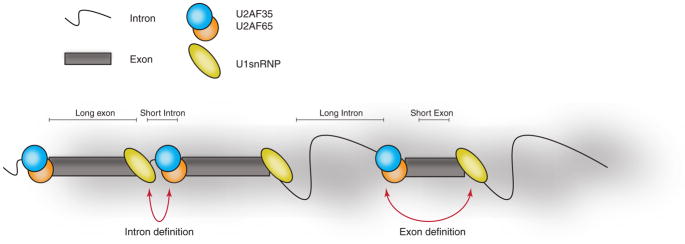
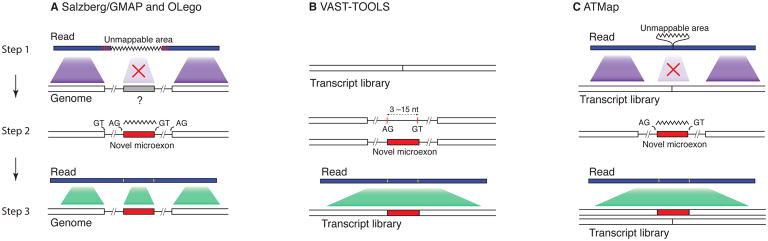
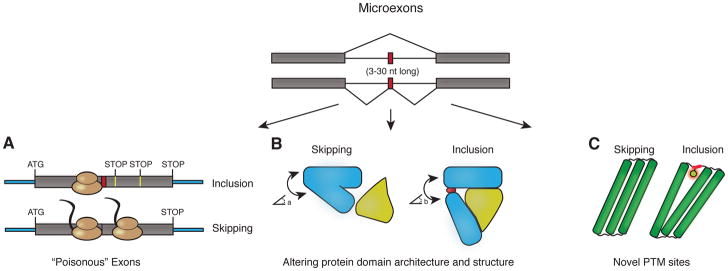
References
-
- Pan Q, Shai O, Lee LJ, Frey BJ, Blencowe BJ. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat Genet. 2008;40:1413–1415. - PubMed
-
- Ule J, Darnell RB. RNA binding proteins and the regulation of neuronal synaptic plasticity. Curr Opin Neurobiol. 2006;16:102–110. - PubMed
-
- Raj B, Blencowe BJ. Alternative splicing in the mammalian nervous system: recent insights into mechanisms and functional roles. Neuron. 2015;87:14–27. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
