

The Macronuclear Genome of Stentor coeruleus Reveals Tiny Introns in a Giant Cell
- PMID: 28190732
- PMCID: PMC5659724
- DOI: 10.1016/j.cub.2016.12.057
The Macronuclear Genome of Stentor coeruleus Reveals Tiny Introns in a Giant Cell
Abstract
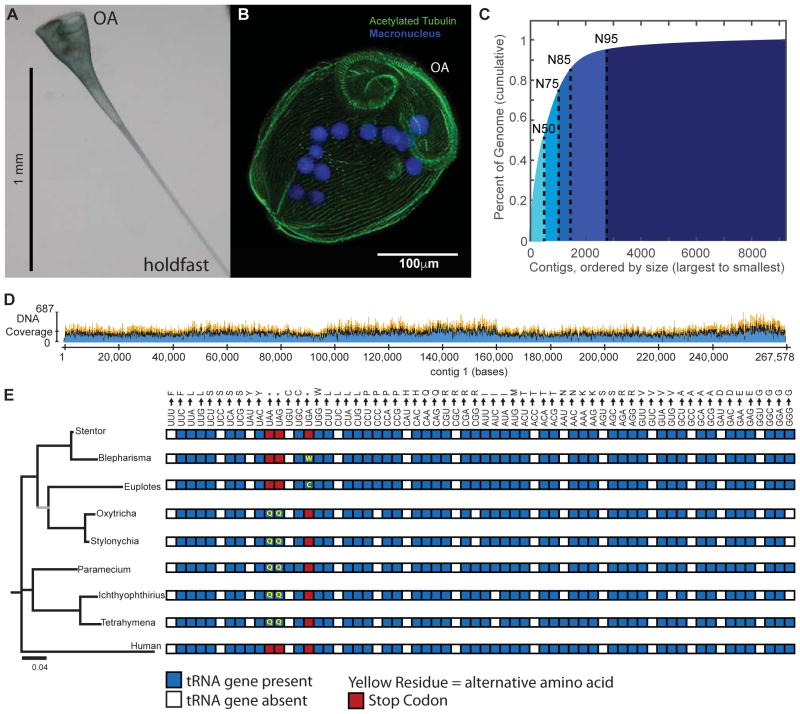
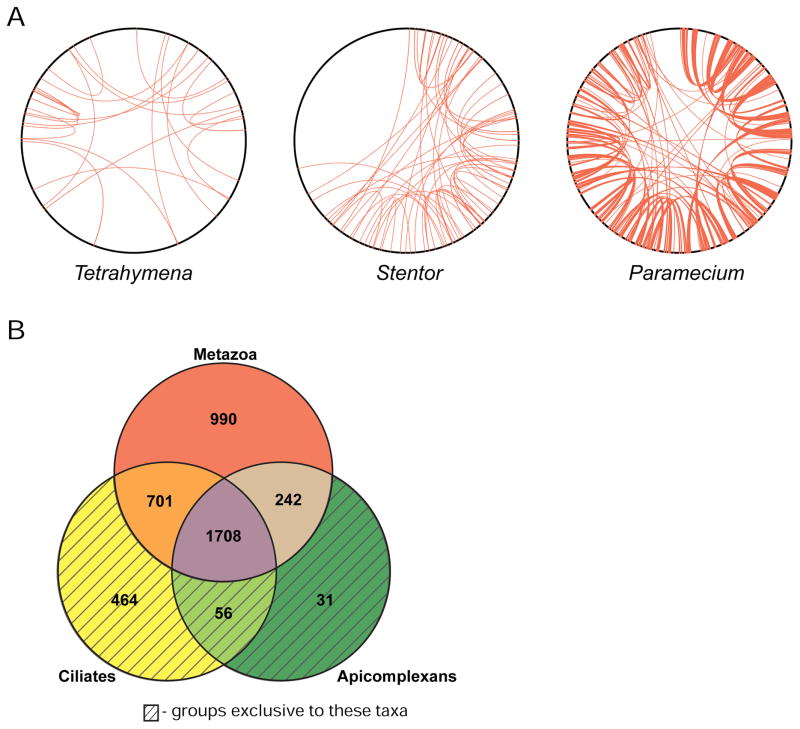
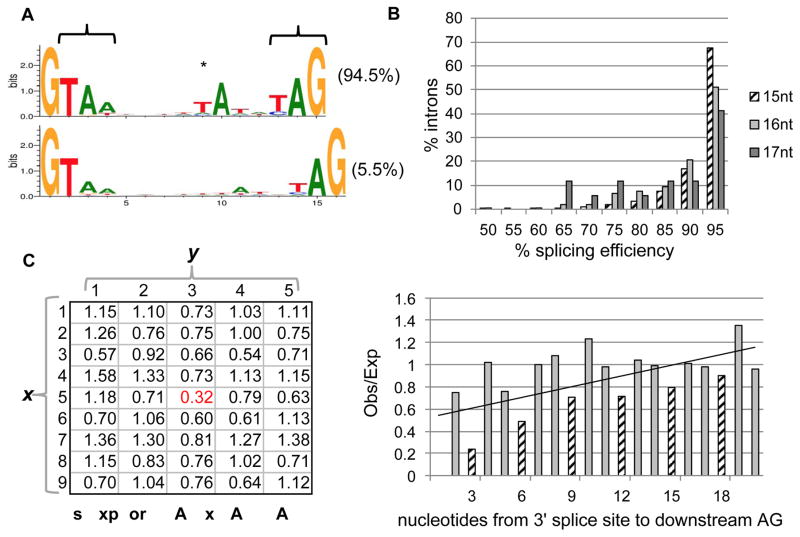
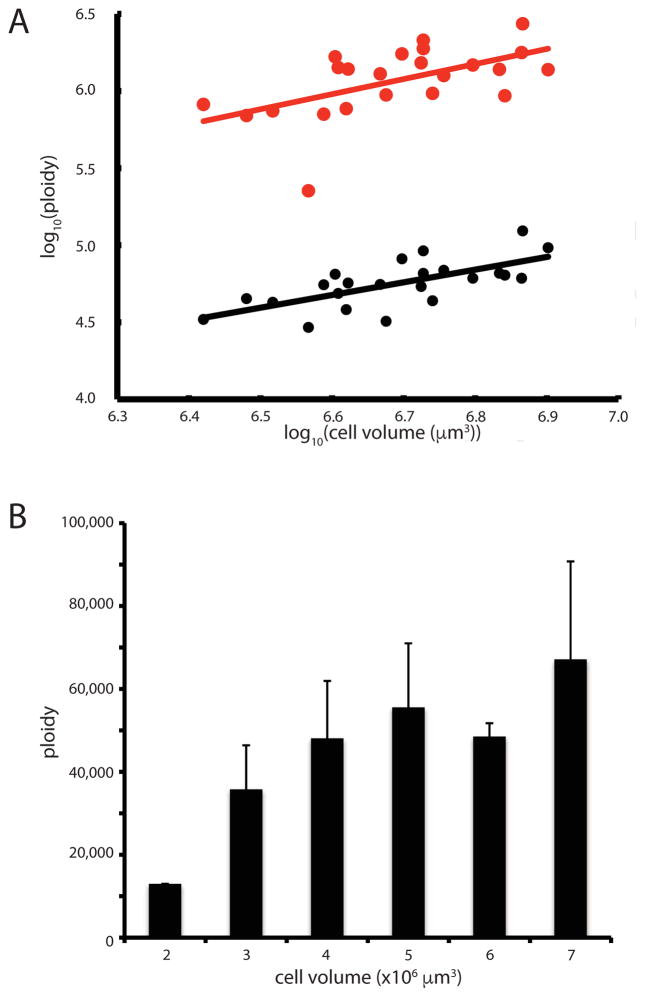
The giant, single-celled organism Stentor coeruleus has a long history as a model system for studying pattern formation and regeneration in single cells. Stentor [1, 2] is a heterotrichous ciliate distantly related to familiar ciliate models, such as Tetrahymena or Paramecium. The primary distinguishing feature of Stentor is its incredible size: a single cell is 1 mm long. Early developmental biologists, including T.H. Morgan [3], were attracted to the system because of its regenerative abilities-if large portions of a cell are surgically removed, the remnant reorganizes into a normal-looking but smaller cell with correct proportionality [2, 3]. These biologists were also drawn to Stentor because it exhibits a rich repertoire of behaviors, including light avoidance, mechanosensitive contraction, food selection, and even the ability to habituate to touch, a simple form of learning usually seen in higher organisms [4]. While early microsurgical approaches demonstrated a startling array of regenerative and morphogenetic processes in this single-celled organism, Stentor was never developed as a molecular model system. We report the sequencing of the Stentor coeruleus macronuclear genome and reveal key features of the genome. First, we find that Stentor uses the standard genetic code, suggesting that ciliate-specific genetic codes arose after Stentor branched from other ciliates. We also discover that ploidy correlates with Stentor's cell size. Finally, in the Stentor genome, we discover the smallest spliceosomal introns reported for any species. The sequenced genome opens the door to molecular analysis of single-cell regeneration in Stentor.
Keywords: U2 snRNA; cell size; ciliate; genetic code; heterotrichidae; intron evolution; macronucleus; ploidy; regeneration; splicing.
Copyright © 2017 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
Comment in
-
Genomics: Stentor's Trumpet Sounds Anew.Curr Biol. 2017 Feb 20;27(4):R146-R148. doi: 10.1016/j.cub.2016.12.059. Curr Biol. 2017. PMID: 28222292
References
-
- Tartar V. The biology of Stentor. New York: Pergammon Press; 1961.
-
- Morgan TH. Regeneration of proportionate structures in Stentor. The Biol Bull. 1901;2:311–328.
-
- Wood DC. Parametric studies of the response decrement produced by mechanical stimuli in the protozoan, Stentor coeruleus. J Neurobiol. 1969;1:345–60. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
