

Species dependent impact of helminth-derived antigens on human macrophages infected with Mycobacterium tuberculosis: Direct effect on the innate anti-mycobacterial response
- PMID: 28192437
- PMCID: PMC5325601
- DOI: 10.1371/journal.pntd.0005390
Species dependent impact of helminth-derived antigens on human macrophages infected with Mycobacterium tuberculosis: Direct effect on the innate anti-mycobacterial response
Abstract
Background: In countries with a high prevalence of tuberculosis there is high coincident of helminth infections that might worsen disease outcome. While Mycobacterium tuberculosis (Mtb) gives rise to a pro-inflammatory Th1 response, a Th2 response is typical of helminth infections. A strong Th2 response has been associated with decreased protection against tuberculosis.
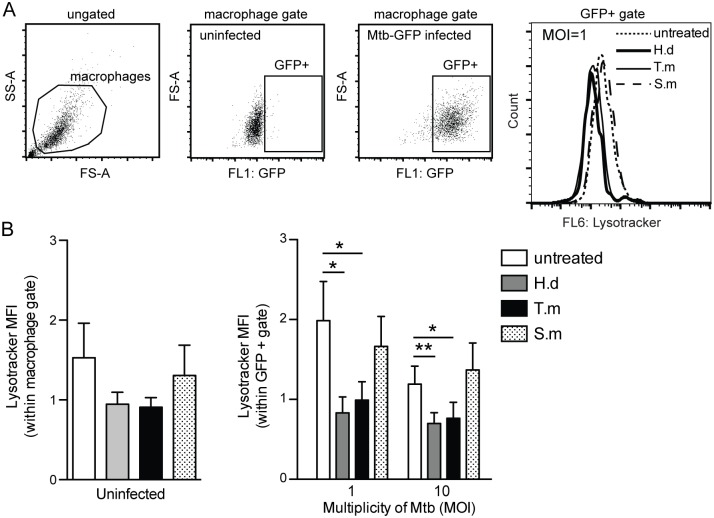
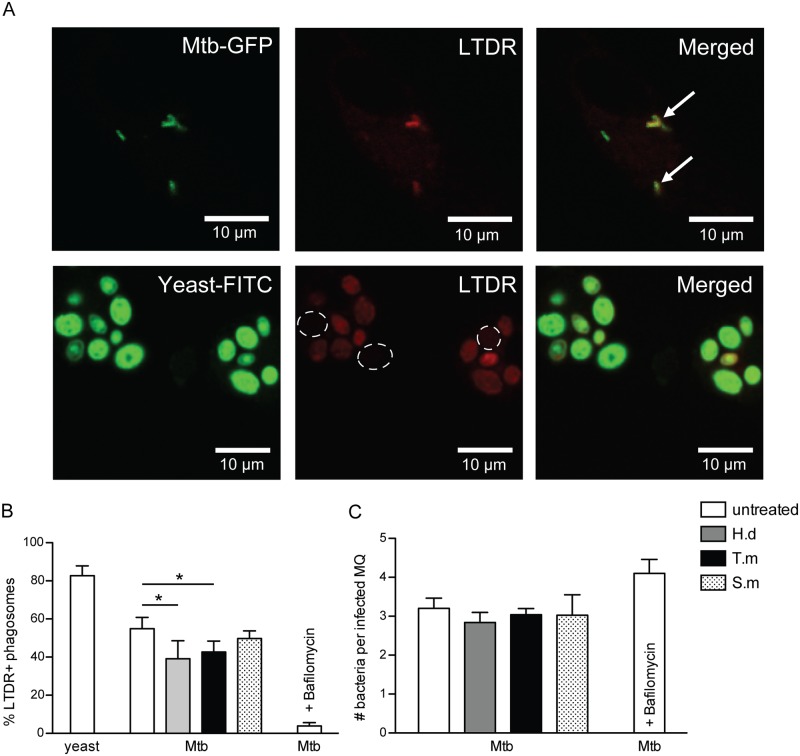
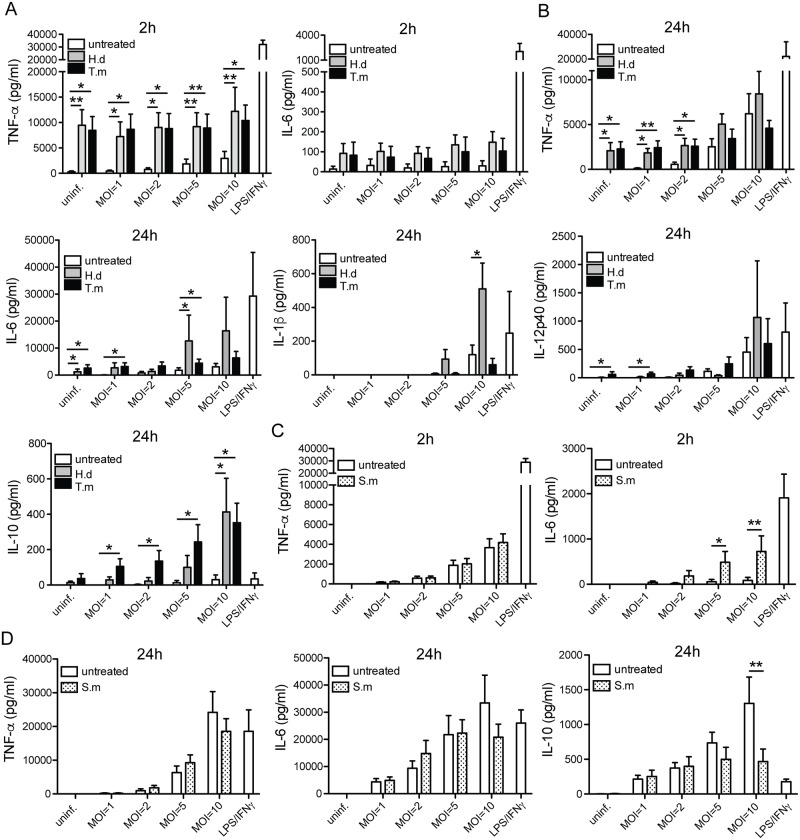
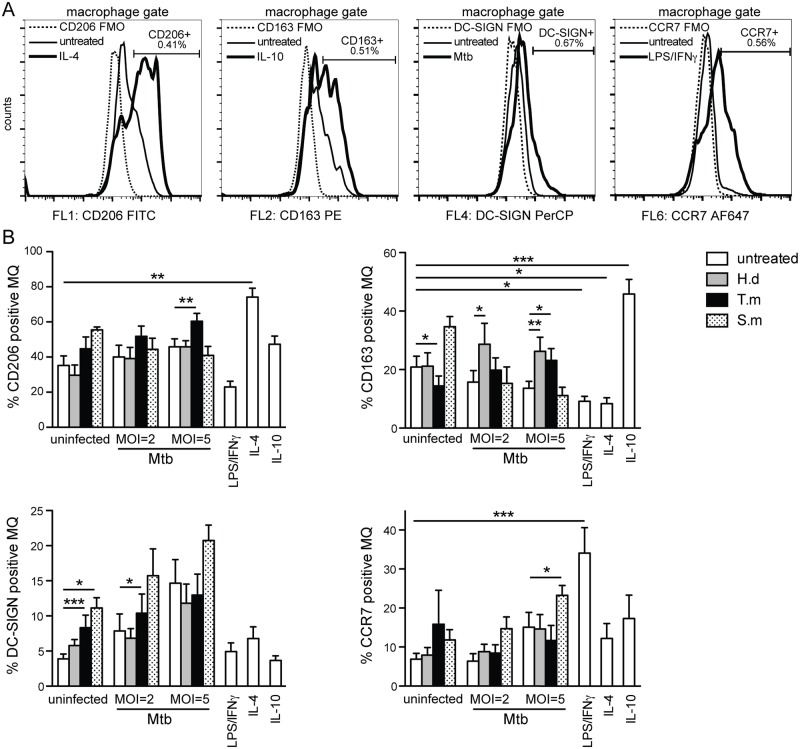
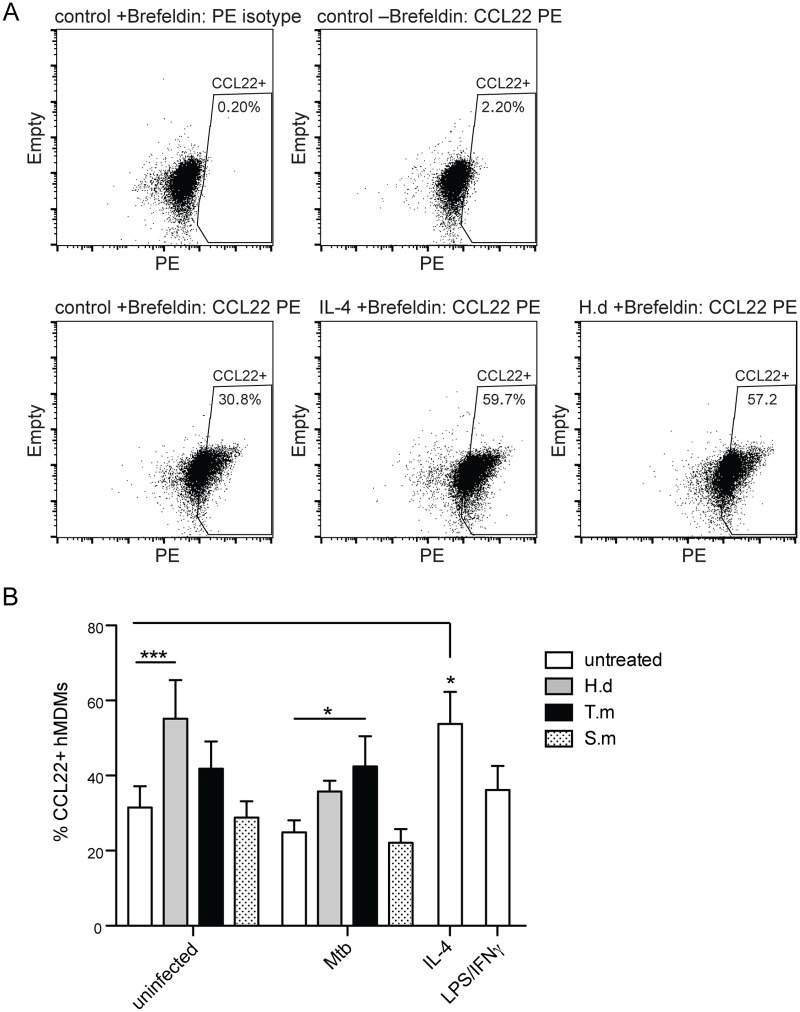
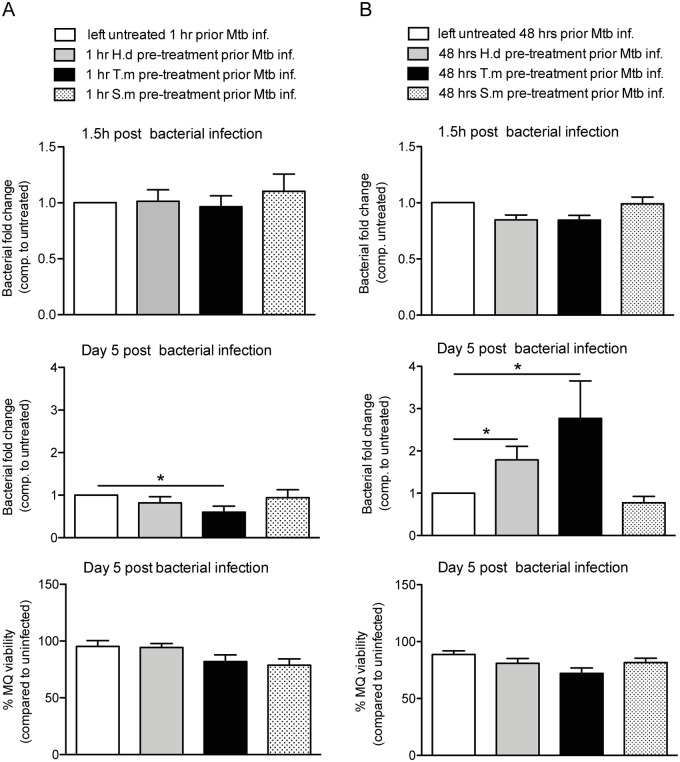
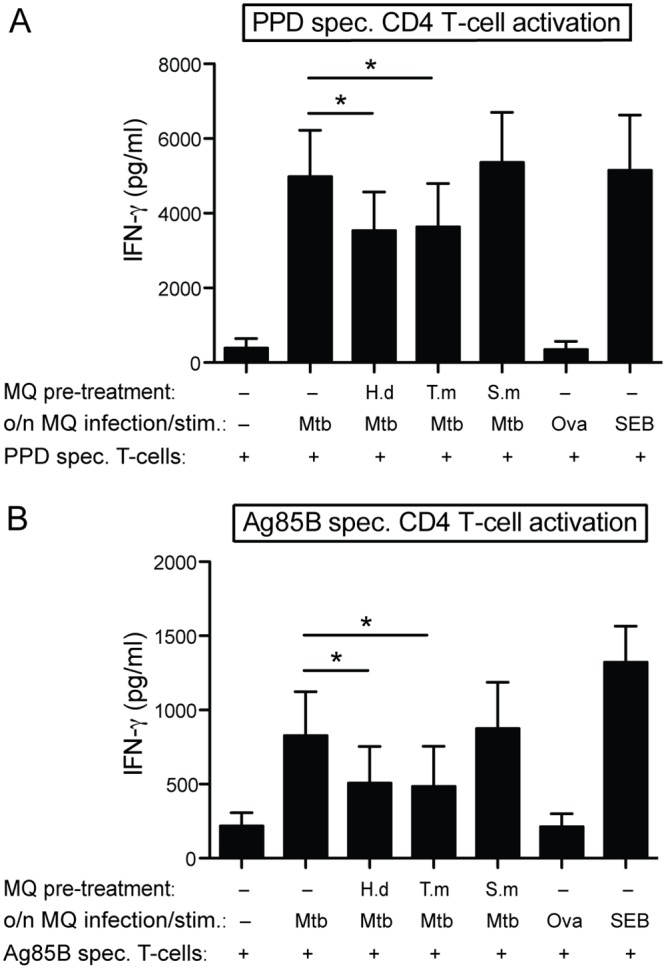
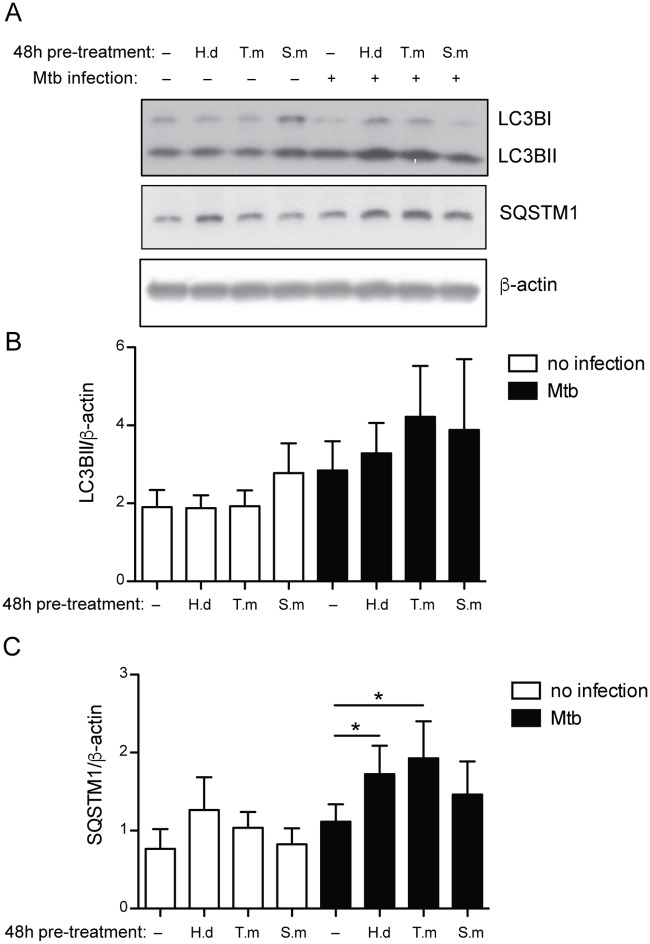
Principal findings: We investigated the direct effect of helminth-derived antigens on human macrophages, hypothesizing that helminths would render macrophages less capable of controlling Mtb. Measuring cytokine output, macrophage surface markers with flow cytometry, and assessing bacterial replication and phagosomal maturation revealed that antigens from different species of helminth directly affect macrophage responses to Mtb. Antigens from the tapeworm Hymenolepis diminuta and the nematode Trichuris muris caused an anti-inflammatory response with M2-type polarization, reduced macrophage phagosome maturation and ability to activate T cells, along with increased Mtb burden, especially in T. muris exposed cells which also induced the highest IL-10 production upon co-infection. However, antigens from the trematode Schistosoma mansoni had the opposite effect causing a decrease in IL-10 production, M1-type polarization and increased control of Mtb.
Conclusion: We conclude that, independent of any adaptive immune response, infection with helminth parasites, in a species-specific manner can influence the outcome of tuberculosis by either enhancing or diminishing the bactericidal function of macrophages.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
