

Metabolic gatekeeper function of B-lymphoid transcription factors
- PMID: 28192788
- PMCID: PMC5621518
- DOI: 10.1038/nature21076
Metabolic gatekeeper function of B-lymphoid transcription factors
Erratum in
-
Author Correction: Metabolic gatekeeper function of B-lymphoid transcription factors.Nature. 2018 Jun;558(7711):E5. doi: 10.1038/s41586-018-0164-5. Nature. 2018. PMID: 29849140
Abstract
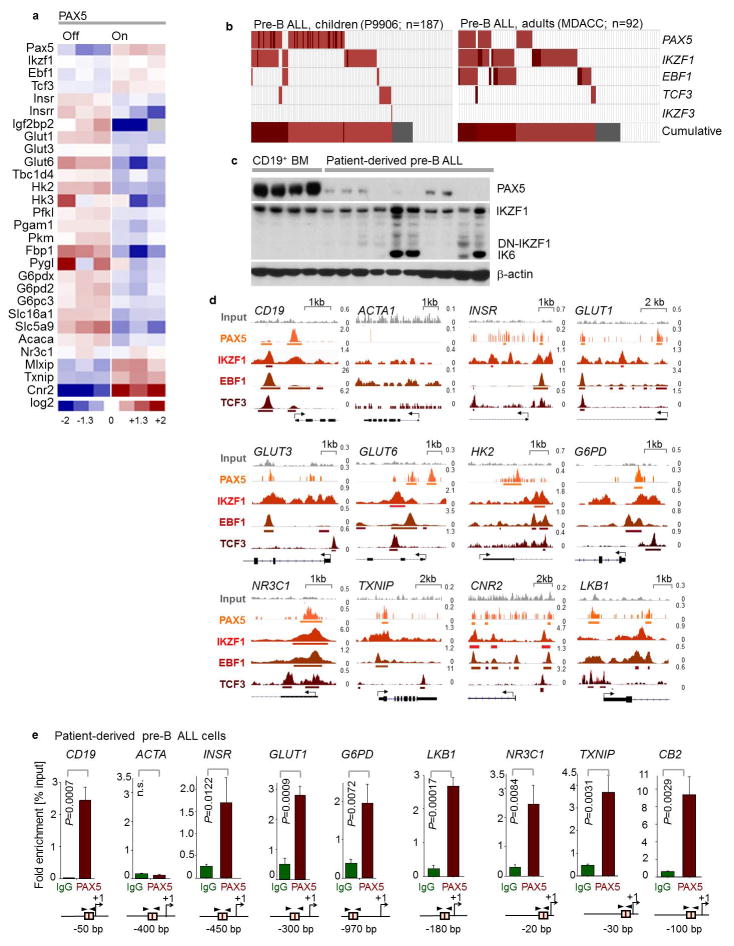
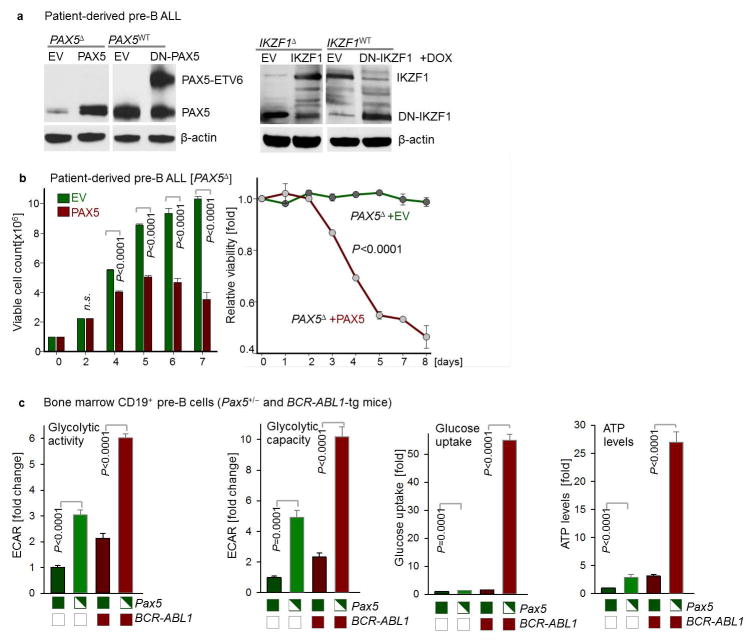
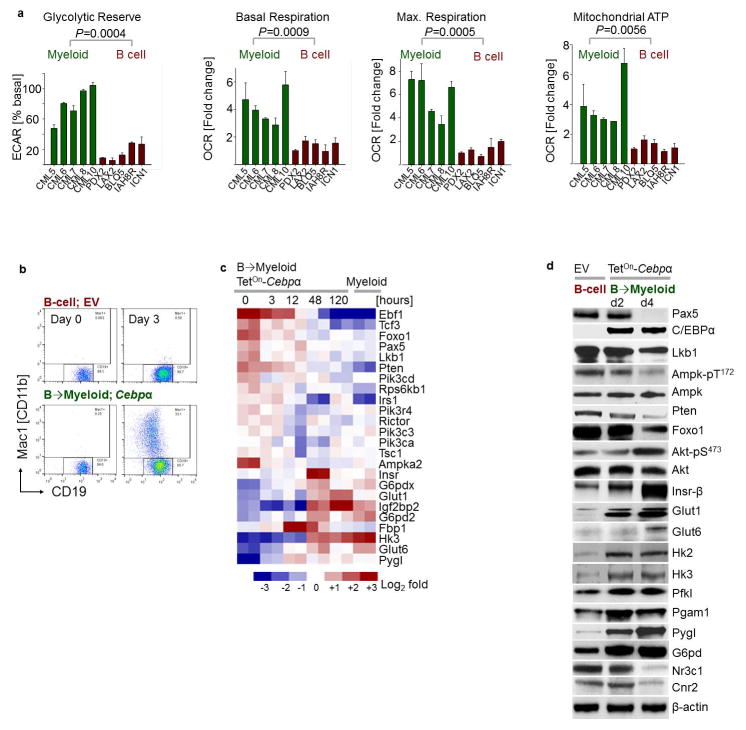
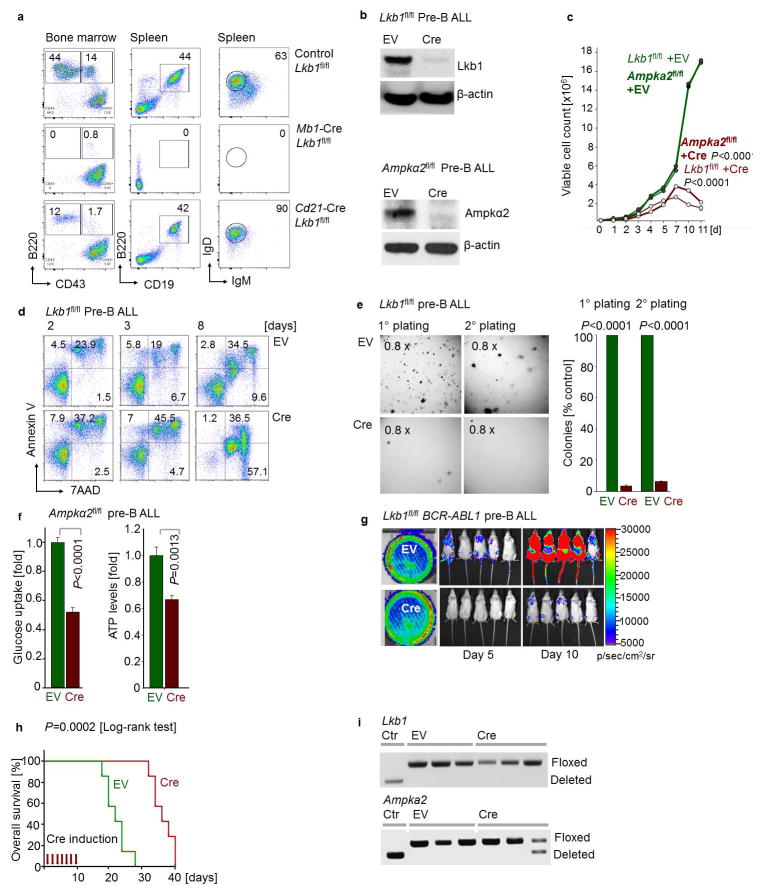
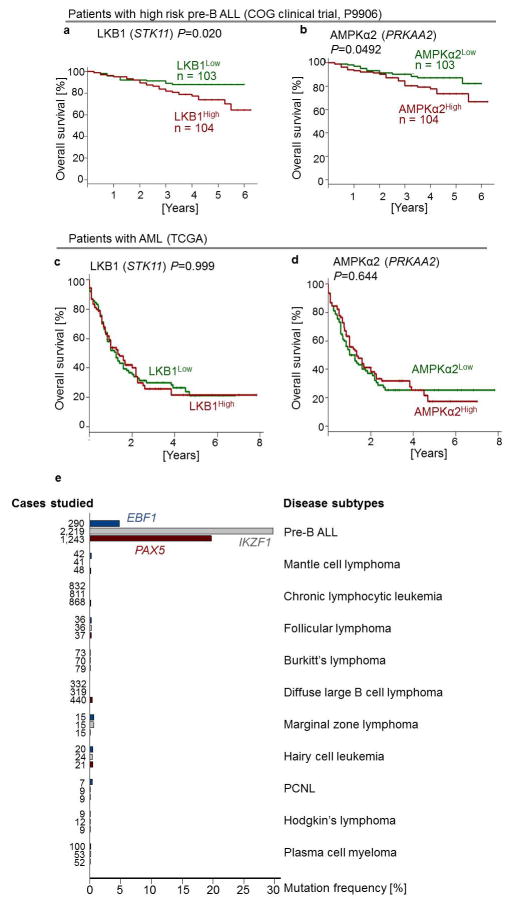
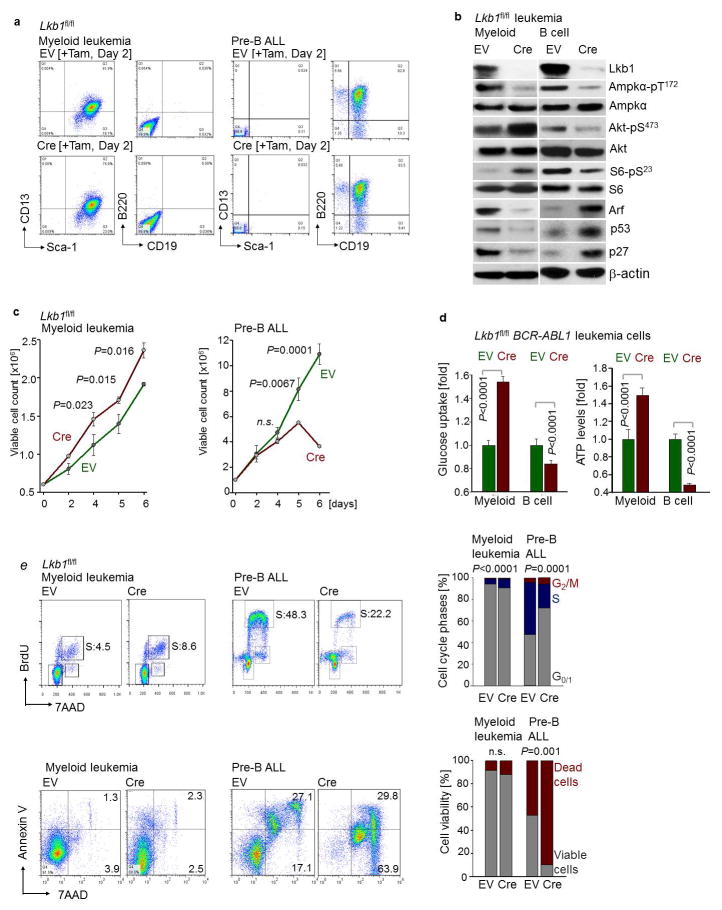
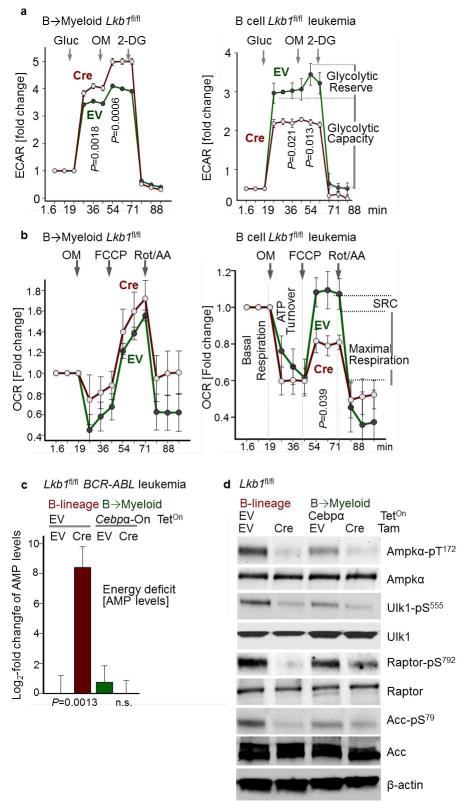
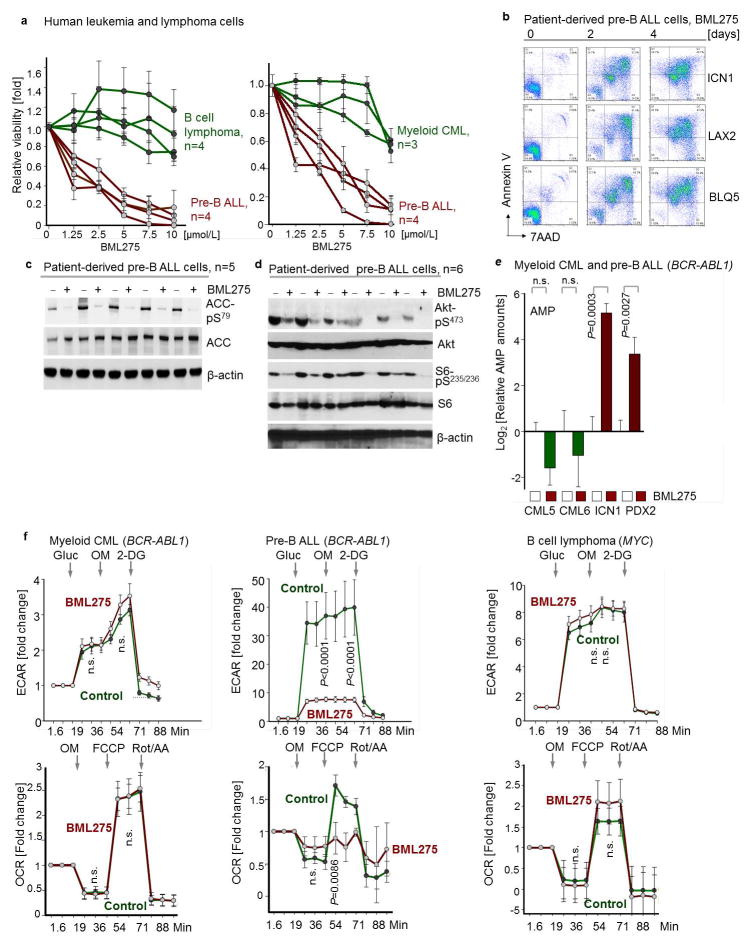
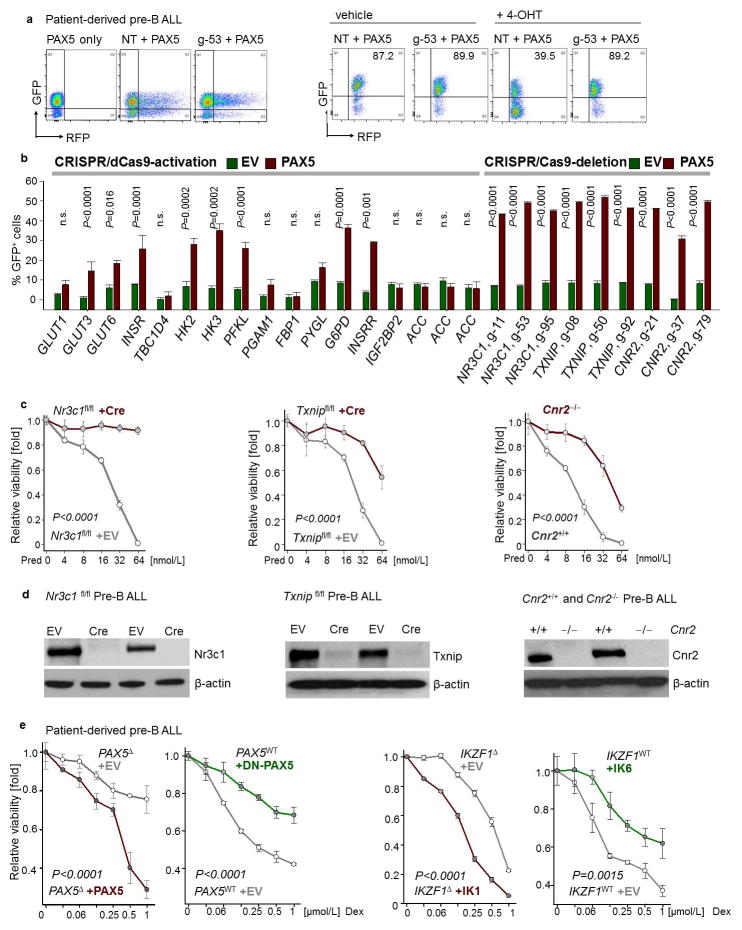
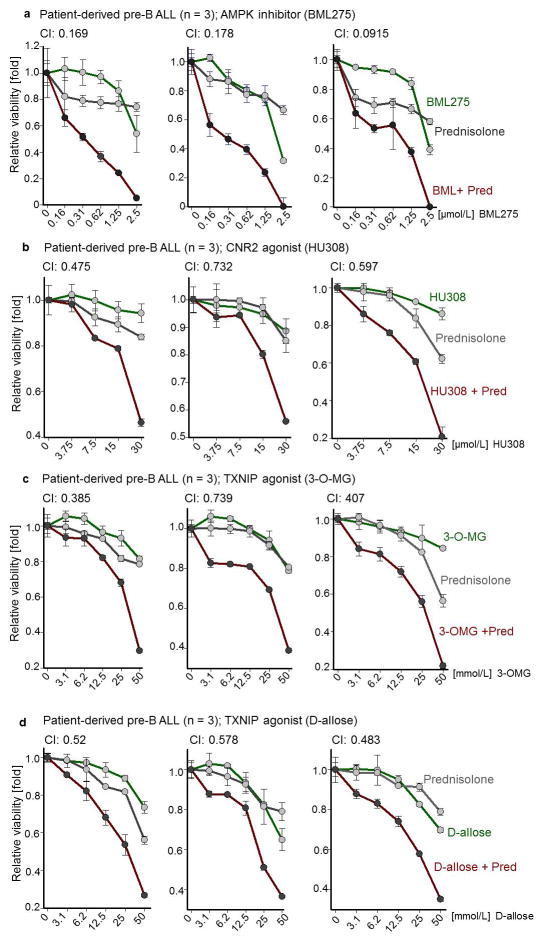
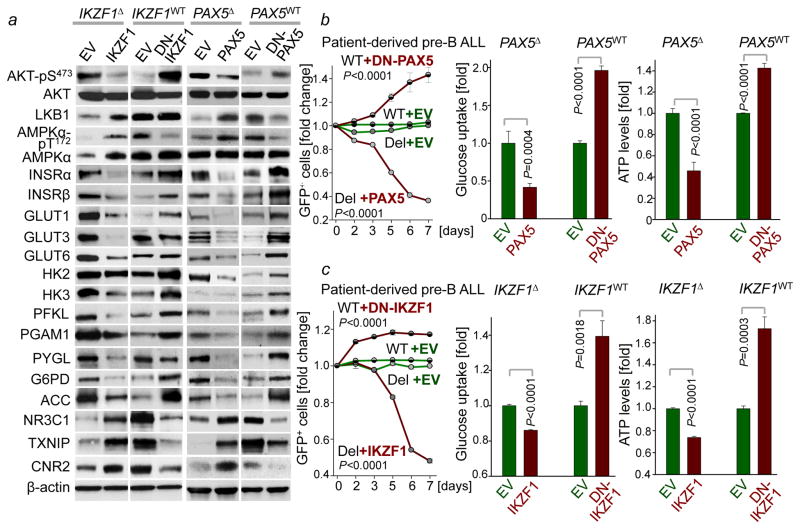
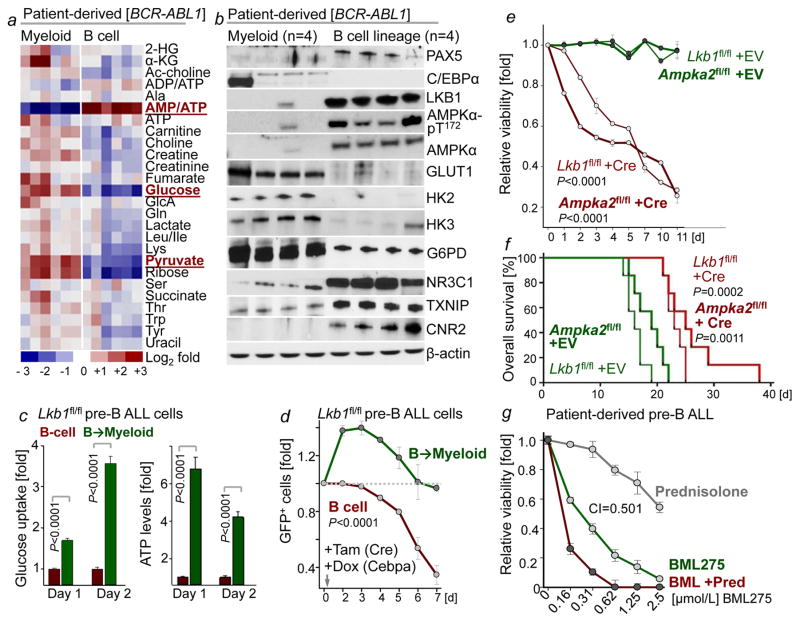
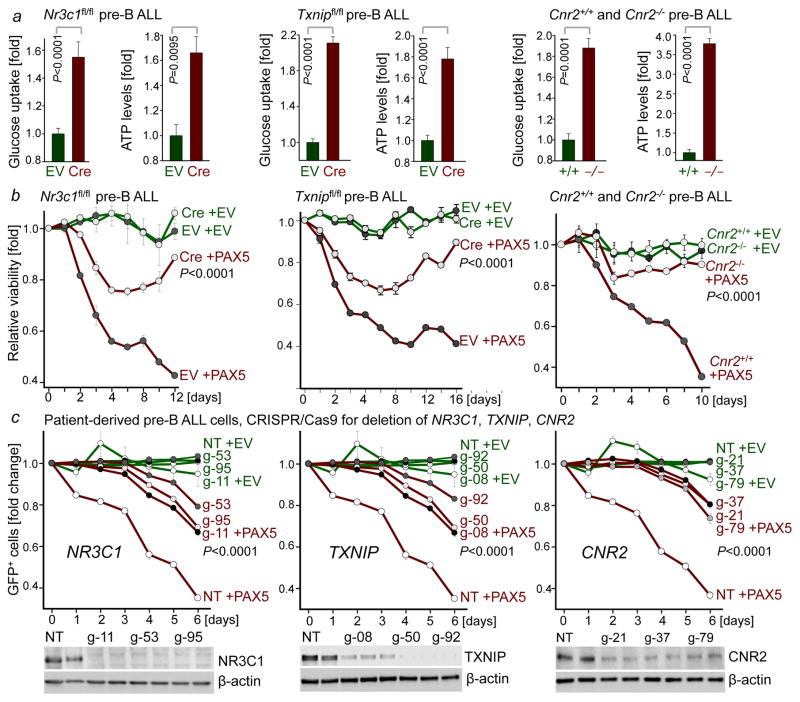
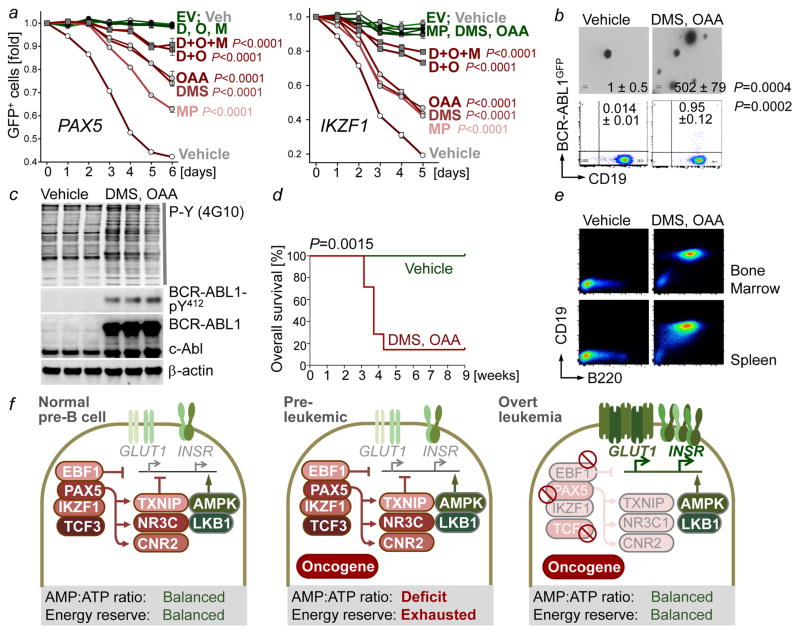
B-lymphoid transcription factors, such as PAX5 and IKZF1, are critical for early B-cell development, yet lesions of the genes encoding these transcription factors occur in over 80% of cases of pre-B-cell acute lymphoblastic leukaemia (ALL). The importance of these lesions in ALL has, until now, remained unclear. Here, by combining studies using chromatin immunoprecipitation with sequencing and RNA sequencing, we identify a novel B-lymphoid program for transcriptional repression of glucose and energy supply. Our metabolic analyses revealed that PAX5 and IKZF1 enforce a state of chronic energy deprivation, resulting in constitutive activation of the energy-stress sensor AMPK. Dominant-negative mutants of PAX5 and IKZF1, however, relieved this glucose and energy restriction. In a transgenic pre-B ALL mouse model, the heterozygous deletion of Pax5 increased glucose uptake and ATP levels by more than 25-fold. Reconstitution of PAX5 and IKZF1 in samples from patients with pre-B ALL restored a non-permissive state and induced energy crisis and cell death. A CRISPR/Cas9-based screen of PAX5 and IKZF1 transcriptional targets identified the products of NR3C1 (encoding the glucocorticoid receptor), TXNIP (encoding a glucose-feedback sensor) and CNR2 (encoding a cannabinoid receptor) as central effectors of B-lymphoid restriction of glucose and energy supply. Notably, transport-independent lipophilic methyl-conjugates of pyruvate and tricarboxylic acid cycle metabolites bypassed the gatekeeper function of PAX5 and IKZF1 and readily enabled leukaemic transformation. Conversely, pharmacological TXNIP and CNR2 agonists and a small-molecule AMPK inhibitor strongly synergized with glucocorticoids, identifying TXNIP, CNR2 and AMPK as potential therapeutic targets. Furthermore, our results provide a mechanistic explanation for the empirical finding that glucocorticoids are effective in the treatment of B-lymphoid but not myeloid malignancies. Thus, B-lymphoid transcription factors function as metabolic gatekeepers by limiting the amount of cellular ATP to levels that are insufficient for malignant transformation.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
Immunometabolism: Powering down leukaemia.Nat Rev Immunol. 2017 Feb 27;17(3):145. doi: 10.1038/nri.2017.17. Nat Rev Immunol. 2017. PMID: 28239166 No abstract available.
References
-
- Nutt SL, Heavey B, Rolink AG, Busslinger M. Commitment to the B-lymphoid lineage depends on the transcription factor Pax5. Nature. 1999;401:556–562. - PubMed
-
- Georgopoulos K, Bigby M, Wang JH, Molnar A, Wu P, Winandy S, Sharpe A. The Ikaros gene is required for the development of all lymphoid lineages. Cell. 1994;79:143–156. - PubMed
-
- Mullighan CG, et al. Genome-wide analysis of genetic alterations in acute lymphoblastic leukaemia. Nature. 2007;446:758–764. - PubMed
-
- Mullighan CG, et al. BCR-ABL1 lymphoblastic leukaemia is characterized by the deletion of Ikaros. Nature. 2008;453:110–114. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R00 CA149088/CA/NCI NIH HHS/United States
- P30 DK063720/DK/NIDDK NIH HHS/United States
- MC_UU_00016/6/MRC_/Medical Research Council/United Kingdom
- Wellcome Trust Senior Investigator Award/International
- R01 CA213138/CA/NCI NIH HHS/United States
- R01 CA157644/CA/NCI NIH HHS/United States
- R01 CA172558/CA/NCI NIH HHS/United States
- R01 CA169458/CA/NCI NIH HHS/United States
- R35 CA197628/CA/NCI NIH HHS/United States
- 55108547/HHMI/Howard Hughes Medical Institute/United States
- R01 CA139032/CA/NCI NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- MC_UU_12009/6/MRC_/Medical Research Council/United Kingdom
- R01 CA137060/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
