

Unresponsive Choline Transporter as a Trait Neuromarker and a Causal Mediator of Bottom-Up Attentional Biases
- PMID: 28193693
- PMCID: PMC5354335
- DOI: 10.1523/JNEUROSCI.3499-16.2017
Unresponsive Choline Transporter as a Trait Neuromarker and a Causal Mediator of Bottom-Up Attentional Biases
Abstract
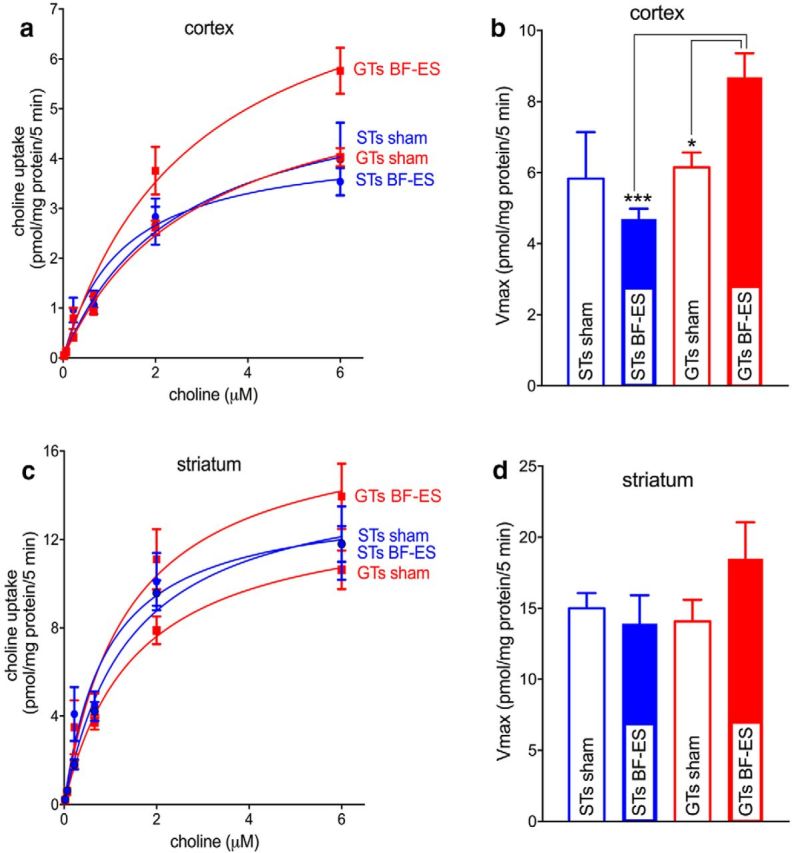
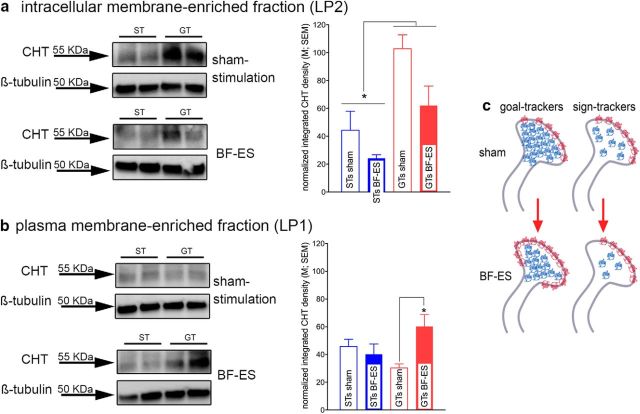
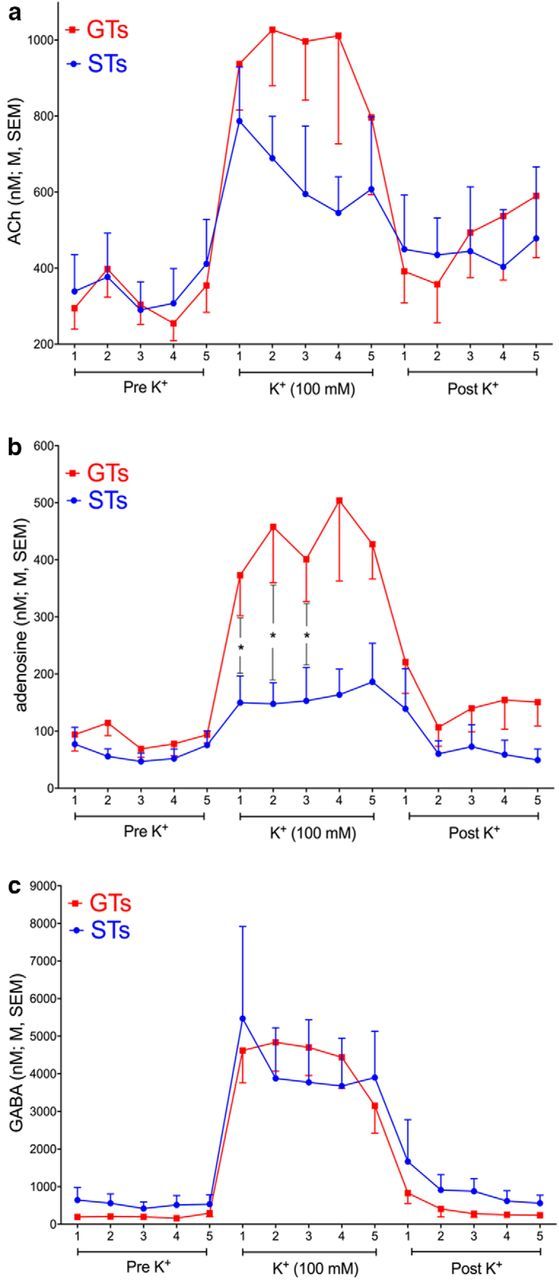
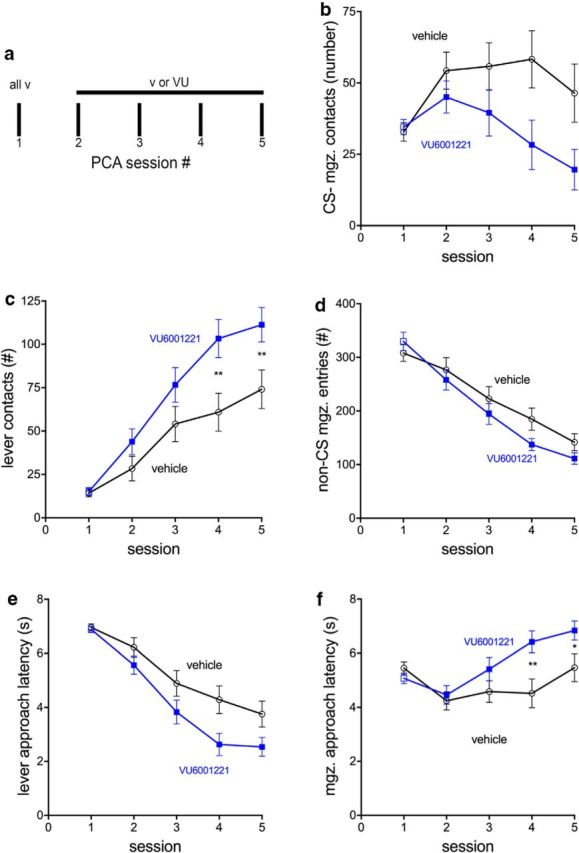
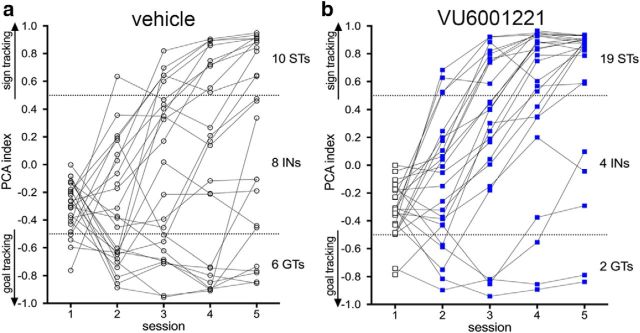
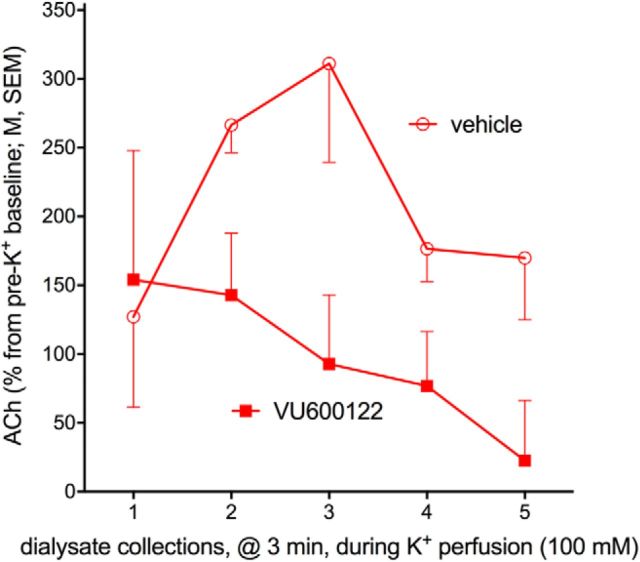
Some rats [sign-trackers (STs)] are prone to attribute incentive salience to reward cues, which can manifest as a propensity to approach and contact pavlovian cues, and for addiction-like behavior. STs also exhibit poor attentional performance, relative to goal-trackers (GTs), which is associated with attenuated acetylcholine (ACh) levels in prefrontal cortex (Paolone et al., 2013). Here, we demonstrate a cellular mechanism, linked to ACh synthesis, that accounts for attenuated cholinergic capacity in STs. First, we found that electrical stimulation of the basal forebrain increased cortical choline transporter (CHT)-mediated choline transport in GTs, paralleled by a redistribution of CHTs to the synaptic plasma membrane. Neither increases in choline uptake nor translocation of CHTs occurred in STs. Second, and consistent with uptake/translocation alterations, STs demonstrated a reduced ability to support cortical ACh release in vivo compared with GTs after reverse-dialysis to elevate extracellular potassium levels. Third, rats were significantly more likely to develop sign-tracking behavior if treated systemically before pavlovian conditioned approach training with the CHT inhibitor VU6001221. Consistent with its proposed mechanisms, administration of VU6001221 attenuated potassium-evoked ACh levels in prefrontal cortex measured with in vivo microdialysis. We propose that loss of CHT-dependent activation of cortical cholinergic activity in STs degrades top-down executive control over behavior, producing a bias for bottom-up or stimulus-driven attention. Such an attentional bias contributes to nonadaptive reward processing and thus identifies a novel mechanism that can support psychopathology, including addiction.SIGNIFICANCE STATEMENT The vulnerability for addiction-like behavior has been associated with psychological traits, such as the propensity to attribute incentive salience to reward cues that is modeled in rats by sign-tracking behavior. Sign-trackers tend to approach and contact cues associated with reward, whereas their counterparts, the goal-trackers, have a preference for approaching the location of the reward. Here, we show that the capacity of presynaptic cholinergic synapses to respond to stimulation by elevating presynaptic choline uptake and releasing acetylcholine is attenuated in sign-trackers. Furthermore, pharmacological inhibition of choline transport induced sign-tracking behavior. Our findings suggest that reduced levels of cholinergic neuromodulation can mediate an attentional bias toward reward-related cues, thereby allowing such cues to exert relatively greater control over behavior.
Keywords: acetylcholine; addiction; attention; choline transporter; motivation; sign-tracking.
Copyright © 2017 the authors 0270-6474/17/372947-13$15.00/0.
Figures
Similar articles
-
Cholinergic control over attention in rats prone to attribute incentive salience to reward cues.J Neurosci. 2013 May 8;33(19):8321-35. doi: 10.1523/JNEUROSCI.0709-13.2013. J Neurosci. 2013. PMID: 23658172 Free PMC article.
-
Diverse Roads to Relapse: A Discriminative Cue Signaling Cocaine Availability Is More Effective in Renewing Cocaine Seeking in Goal Trackers Than Sign Trackers and Depends on Basal Forebrain Cholinergic Activity.J Neurosci. 2017 Jul 26;37(30):7198-7208. doi: 10.1523/JNEUROSCI.0990-17.2017. Epub 2017 Jun 28. J Neurosci. 2017. PMID: 28659281 Free PMC article.
-
Neuro-Immune Modulation of Cholinergic Signaling in an Addiction Vulnerability Trait.eNeuro. 2023 Mar 2;10(3):ENEURO.0023-23.2023. doi: 10.1523/ENEURO.0023-23.2023. Print 2023 Mar. eNeuro. 2023. PMID: 36810148 Free PMC article.
-
Cholinergic genetics of visual attention: Human and mouse choline transporter capacity variants influence distractibility.J Physiol Paris. 2016 Sep;110(1-2):10-18. doi: 10.1016/j.jphysparis.2016.07.001. Epub 2016 Jul 9. J Physiol Paris. 2016. PMID: 27404793 Free PMC article. Review.
-
The choline transporter resurfaces: new roles for synaptic vesicles?Mol Interv. 2004 Feb;4(1):22-37. doi: 10.1124/mi.4.1.22. Mol Interv. 2004. PMID: 14993474 Review.
Cited by
-
The neuroscience of cognitive-motivational styles: Sign- and goal-trackers as animal models.Behav Neurosci. 2018 Feb;132(1):1-12. doi: 10.1037/bne0000226. Epub 2018 Jan 22. Behav Neurosci. 2018. PMID: 29355335 Free PMC article.
-
The paraventricular thalamus is a critical mediator of top-down control of cue-motivated behavior in rats.Elife. 2019 Sep 10;8:e49041. doi: 10.7554/eLife.49041. Elife. 2019. PMID: 31502538 Free PMC article.
-
Cholinergic double duty: cue detection and attentional control.Curr Opin Psychol. 2019 Oct;29:102-107. doi: 10.1016/j.copsyc.2018.12.026. Epub 2019 Jan 4. Curr Opin Psychol. 2019. PMID: 30711909 Free PMC article. Review.
-
Nicotine Enhances Goal-Tracking in Ethanol and Food Pavlovian Conditioned Approach Paradigms.Front Neurosci. 2021 Aug 18;15:561766. doi: 10.3389/fnins.2021.561766. eCollection 2021. Front Neurosci. 2021. PMID: 34483813 Free PMC article.
-
Cortico-striatal action control inherent of opponent cognitive-motivational styles.bioRxiv [Preprint]. 2024 Dec 5:2024.03.12.584623. doi: 10.1101/2024.03.12.584623. bioRxiv. 2024. Update in: Elife. 2025 Feb 19;13:RP100988. doi: 10.7554/eLife.100988. PMID: 38559086 Free PMC article. Updated. Preprint.
References
-
- Apparsundaram S, Martinez V, Parikh V, Kozak R, Sarter M (2005) Increased capacity and density of choline transporters situated in synaptic membranes of the right medial prefrontal cortex of attentional task-performing rats. J Neurosci 25:3851–3856. 10.1523/JNEUROSCI.0205-05.2005 - DOI - PMC - PubMed
-
- Bertron JL, Ennis EA, Tarr CJ, Wright J, Dickerson JW, Locuson CW, Blobaum AL, Rook JM, Blakely RD, Lindsley CW (2016) Optimization of the choline transporter (CHT) inhibitor ML352: development of VU6001221, an improved in vivo tool compound. Bioorg Med Chem Lett 26:4637–4640. 10.1016/j.bmcl.2016.08.062 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources