

Experimental Arthritis Is Dependent on Mouse Mast Cell Protease-5
- PMID: 28193842
- PMCID: PMC5392683
- DOI: 10.1074/jbc.M116.773416
Experimental Arthritis Is Dependent on Mouse Mast Cell Protease-5
Abstract
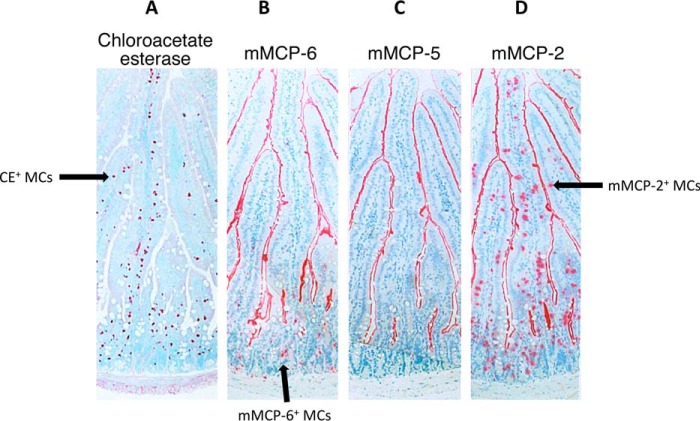
The constitutive heparin+ (HP) mast cells (MCs) in mice express mouse MC protease (mMCP)-5 and carboxypeptidase A (mMC-CPA). The amino acid sequence of mMCP-5 is most similar to that of human chymase-1, as are the nucleotide sequences of their genes and transcripts. Using a homologous recombination approach, a C57BL/6 mouse line was created that possessed a disrupted mMCP-5 gene. The resulting mice were fertile and had no obvious developmental abnormality. Lack of mMCP-5 protein did not alter the granulation of the IL-3/IL-9-dependent mMCP-2+ MCs in the jejunal mucosa of Trichinella spiralis-infected mice. In contrast, the constitutive HP+ MCs in the tongues of mMCP-5-null mice were poorly granulated and lacked mMC-CPA protein. Bone marrow-derived MCs were readily developed from the transgenic mice using IL-3. Although these MCs contained high levels of mMC-CPA mRNA, they also lacked the latter exopeptidase. mMCP-5 protein is therefore needed to target translated mMC-CPA to the secretory granule along with HP-containing serglycin proteoglycans. Alternately, mMCP-5 is needed to protect mMC-CPA from autolysis in the cell's granules. Fibronectin was identified as a target of mMCP-5, and the exocytosis of mMCP-5 from the MCs in the mouse's peritoneal cavity resulted in the expression of metalloproteinase protease-9, which has been implicated in arthritis. In support of the latter finding, experimental arthritis was markedly reduced in mMCP-5-null mice relative to wild-type mice in two disease models.
Keywords: arthritis; carboxypeptidase; heparin; mast cell; serine protease.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures









References
-
- Caughey G. H., Zerweck E. H., and Vanderslice P. (1991) Structure, chromosomal assignment, and deduced amino acid sequence of a human gene for mast cell chymase. J. Biol. Chem. 266, 12956–12963 - PubMed
-
- Urata H., Kinoshita A., Perez D. M., Misono K. S., Bumpus F. M., Graham R. M., and Husain A. (1991) Cloning of the gene and cDNA for human heart chymase. J. Biol. Chem. 266, 17173–17179 - PubMed
-
- Urata H., Kinoshita A., Misono K. S., Bumpus F. M., and Husain A. (1990) Identification of a highly specific chymase as the major angiotensin II-forming enzyme in the human heart. J. Biol. Chem. 265, 22348–22357 - PubMed
-
- Semaan W., Desbiens L., Houde M., Labonté J., Gagnon H., Yamamoto D., Takai S., Laidlaw T., Bkaily G., Schwertani A., Pejler G., Levesque C., Desjardins R., Day R., and D'Orléans-Juste P. (2015) Chymase inhibitor-sensitive synthesis of endothelin-1 (1–31) by recombinant mouse mast cell protease 4 and human chymase. Biochem. Pharmacol. 94, 91–100 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
