

Trade-offs between enzyme fitness and solubility illuminated by deep mutational scanning
- PMID: 28196882
- PMCID: PMC5338495
- DOI: 10.1073/pnas.1614437114
Trade-offs between enzyme fitness and solubility illuminated by deep mutational scanning
Abstract
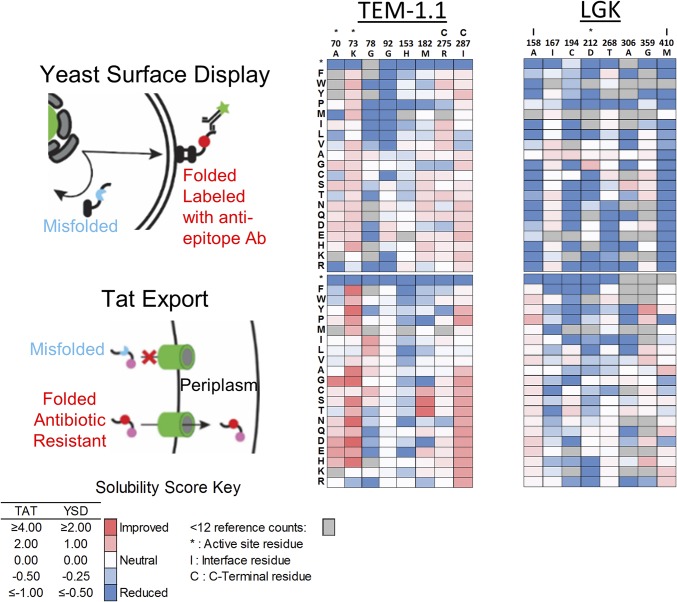
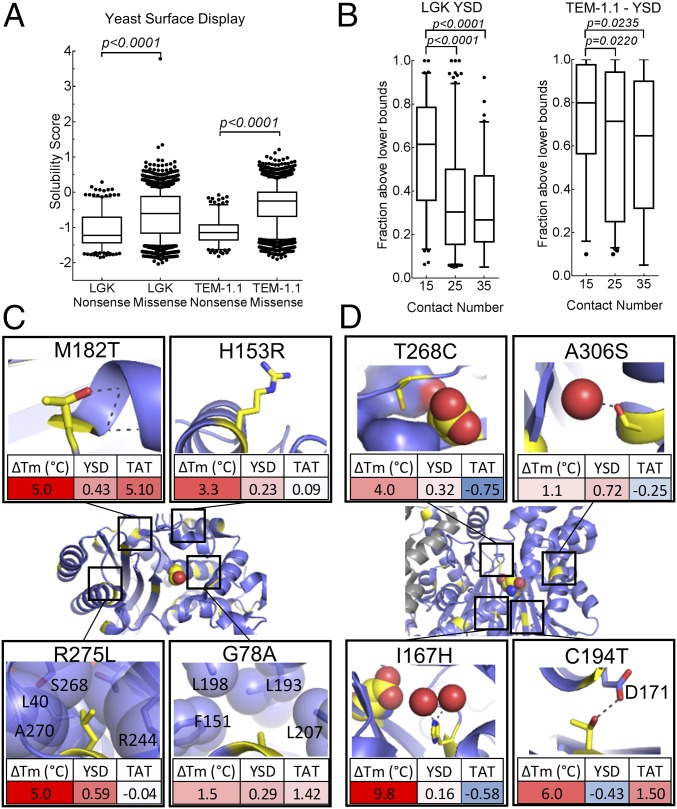
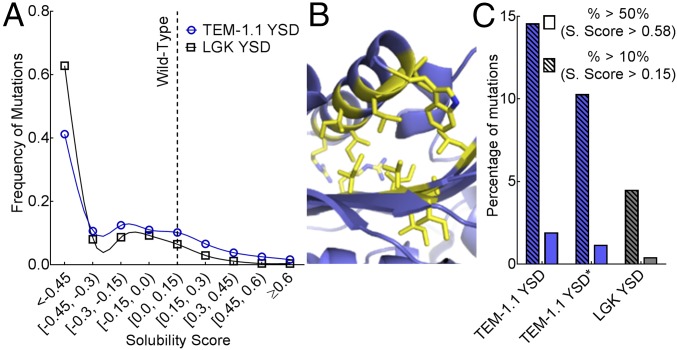
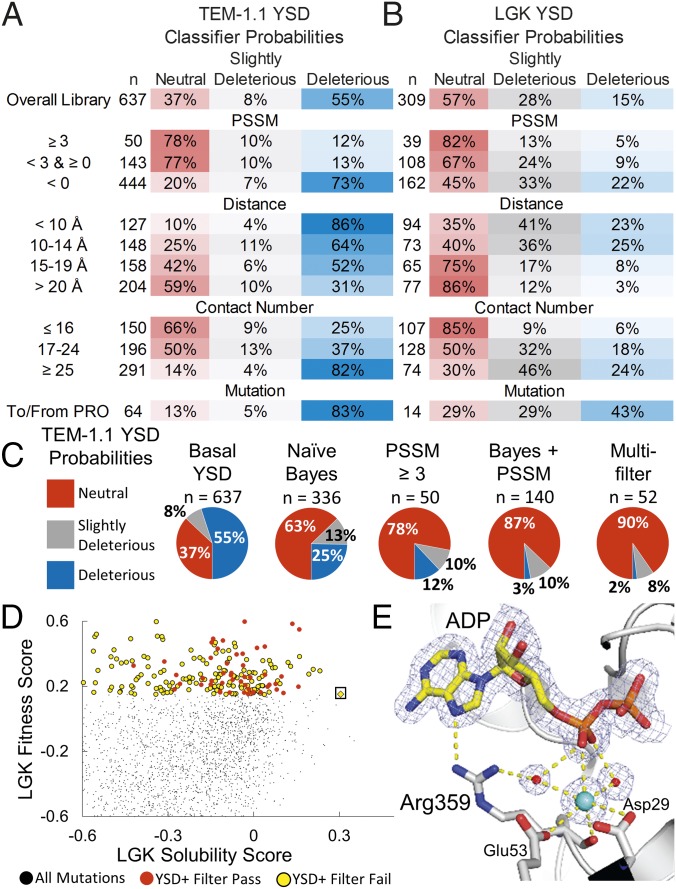
Proteins are marginally stable, and an understanding of the sequence determinants for improved protein solubility is highly desired. For enzymes, it is well known that many mutations that increase protein solubility decrease catalytic activity. These competing effects frustrate efforts to design and engineer stable, active enzymes without laborious high-throughput activity screens. To address the trade-off between enzyme solubility and activity, we performed deep mutational scanning using two different screens/selections that purport to gauge protein solubility for two full-length enzymes. We assayed a TEM-1 beta-lactamase variant and levoglucosan kinase (LGK) using yeast surface display (YSD) screening and a twin-arginine translocation pathway selection. We then compared these scans with published experimental fitness landscapes. Results from the YSD screen could explain 37% of the variance in the fitness landscapes for one enzyme. Five percent to 10% of all single missense mutations improve solubility, matching theoretical predictions of global protein stability. For a given solubility-enhancing mutation, the probability that it would retain wild-type fitness was correlated with evolutionary conservation and distance to active site, and anticorrelated with contact number. Hybrid classification models were developed that could predict solubility-enhancing mutations that maintain wild-type fitness with an accuracy of 90%. The downside of using such classification models is the removal of rare mutations that improve both fitness and solubility. To reveal the biophysical basis of enhanced protein solubility and function, we determined the crystallographic structure of one such LGK mutant. Beyond fundamental insights into trade-offs between stability and activity, these results have potential biotechnological applications.
Keywords: deep mutational scanning; fitness landscapes; high-throughput screening; protein solubility; yeast surface display.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Sormanni P, Aprile FA, Vendruscolo M. The CamSol method of rational design of protein mutants with enhanced solubility. J Mol Biol. 2015;427(2):478–490. - PubMed
-
- Waldo GS. Genetic screens and directed evolution for protein solubility. Curr Opin Chem Biol. 2003;7(1):33–38. - PubMed
-
- Park S, et al. Limitations of yeast surface display in engineering proteins of high thermostability. Protein Eng Des Sel. 2006;19(5):211–217. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
