

Vacuolar ion channels in the liverwort Marchantia polymorpha: influence of ion channel inhibitors
- PMID: 28197715
- PMCID: PMC5391376
- DOI: 10.1007/s00425-017-2661-4
Vacuolar ion channels in the liverwort Marchantia polymorpha: influence of ion channel inhibitors
Abstract
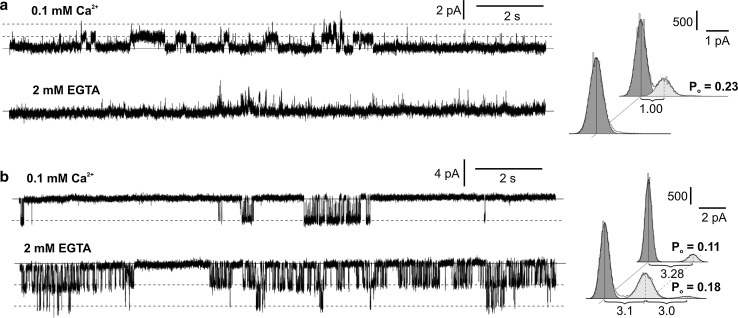
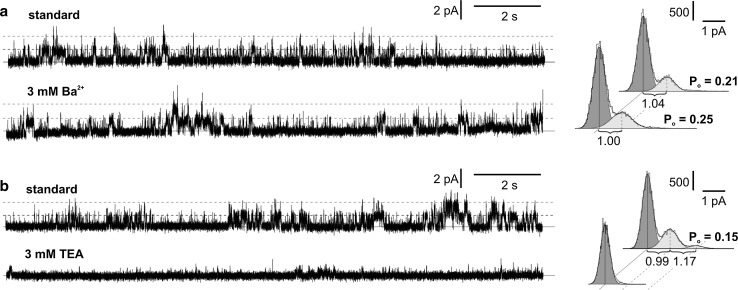
Potassium-permeable slow activating vacuolar channels (SV) and chloride-permeable channels in the vacuole of the liverwort Marchantia polymorpha were characterized in respect to calcium dependence, selectivity, and pharmacology. The patch-clamp method was used in the study of ion channel activity in the vacuoles from the liverwort Marchantia polymorpha. The whole-vacuole recordings allowed simultaneous observation of two types of currents-predominant slow activated currents recorded at positive voltages and fast activated currents recorded at negative voltages. Single-channel recordings carried out in the gradient of KCl indicated that slow activated currents were carried by potassium-permeable slowly activating vacuolar channels (SV) and fast activated currents-by chloride-permeable channels. Both types of the channels were dependent in an opposite way on calcium, since elimination of this ion from the cytoplasmic side caused inhibition of SV channels, but the open probability of chloride-permeable channels even increased. The dependence of the activity of both channels on different types of ion channel inhibitors was studied. SV channels exhibited different sensitivity to potassium channel inhibitors. These channels were insensitive to 3 mM Ba2+, but were blocked by 3 mM tetraethyl ammonium (TEA). Moreover, the activity of the channels was modified in a different way by calcium channel inhibitors. 200 µM Gd3+ was an effective blocker, but 50 µM ruthenium red evoked bursts of the channel activity resulting in an increase in the open probability. Different effectiveness of anion channel inhibitors was observed in chloride-permeable channels. After the application of 100 µM Zn2+, a decrease in the open probability was recorded but the channels were still active. 50 µM DIDS was more effective, as it completely blocked the channels.
Keywords: Calcium dependence; Chloride channels; Patch-clamp; Pharmacology; SV channels; Vacuole.
Figures




References
-
- Allen GJ, Amtmann A, Sanders D. Calcium-dependent and calcium-independent K+ mobilization channels in Vicia faba guard cell vacuoles. J Exp Bot. 1998;49:305–318. doi: 10.1093/jxb/49.Special_Issue.305. - DOI
-
- Bertl A, Blumwald E, Coronado R, Eisenberg R, Findlay G, Gradmann D, Hille B, Kohler K, Kolb HA, Macrobbie E, Meissner G, Miller C, Neher E, Palade P, Pantoja O, Sanders D, Schroeder J, Slayman C, Spanswick R, Walker A, Williams A. Electrical measurements on endomembranes. Science. 1992;258(5084):873–874. doi: 10.1126/science.1439795. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
