

Next-generation transgenic cotton: pyramiding RNAi and Bt counters insect resistance
- PMID: 28199783
- PMCID: PMC5552478
- DOI: 10.1111/pbi.12709
Next-generation transgenic cotton: pyramiding RNAi and Bt counters insect resistance
Abstract
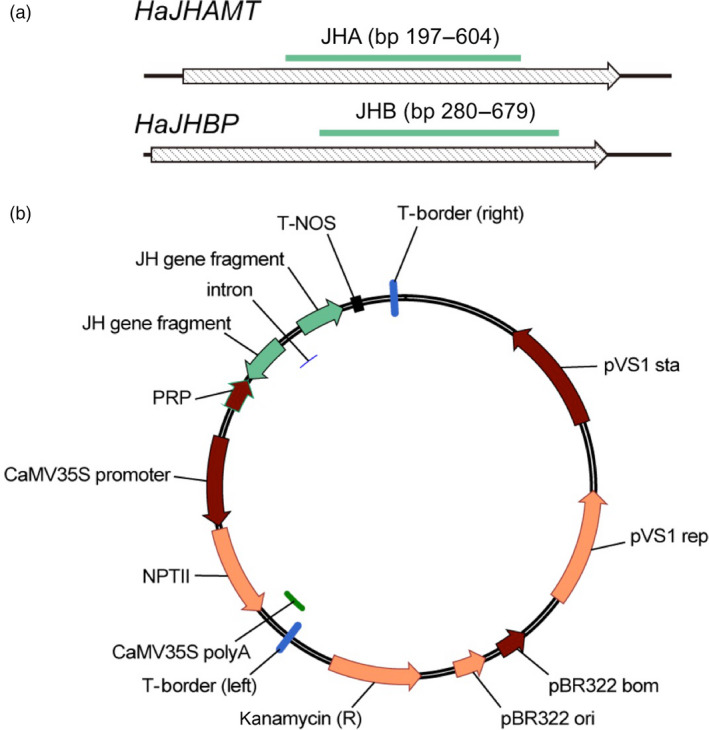
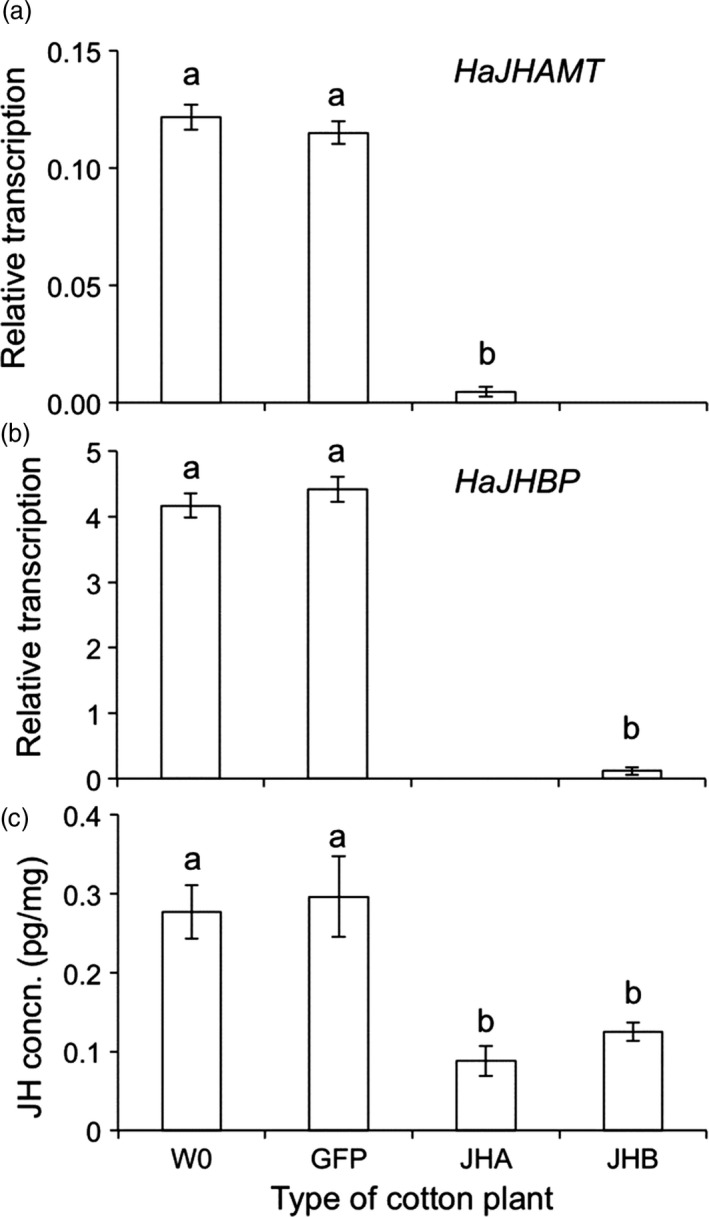
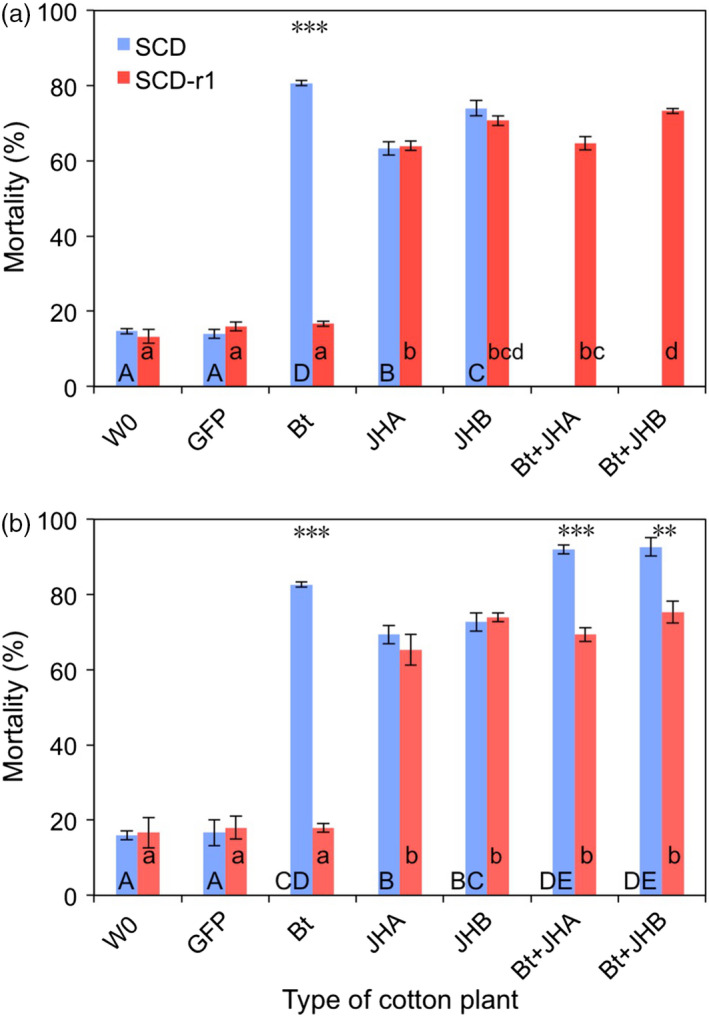
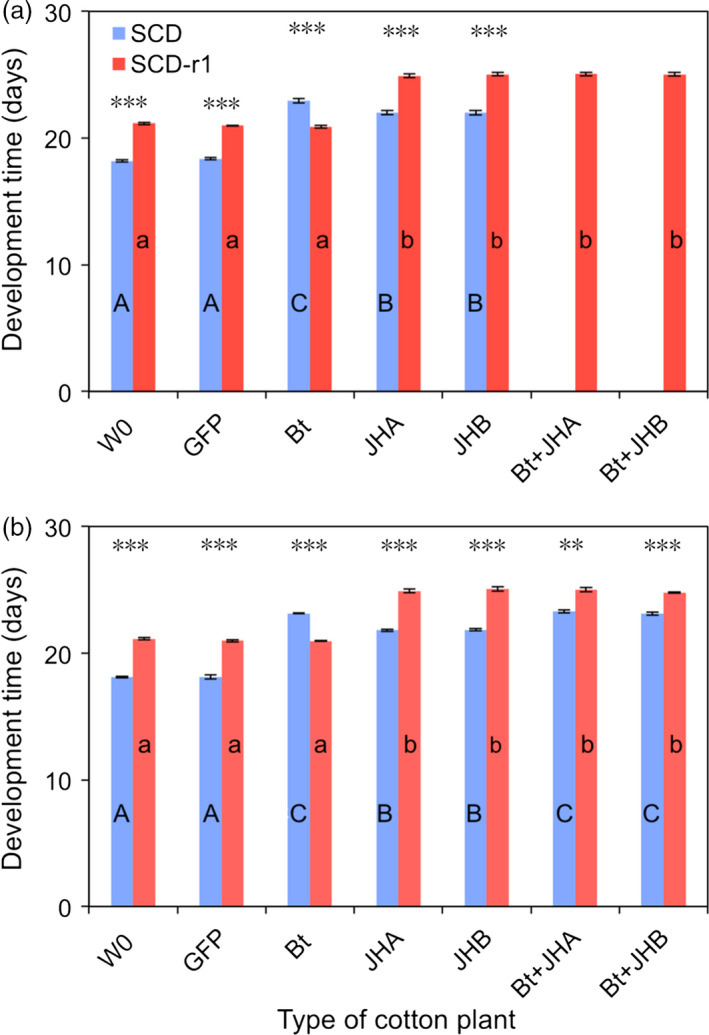
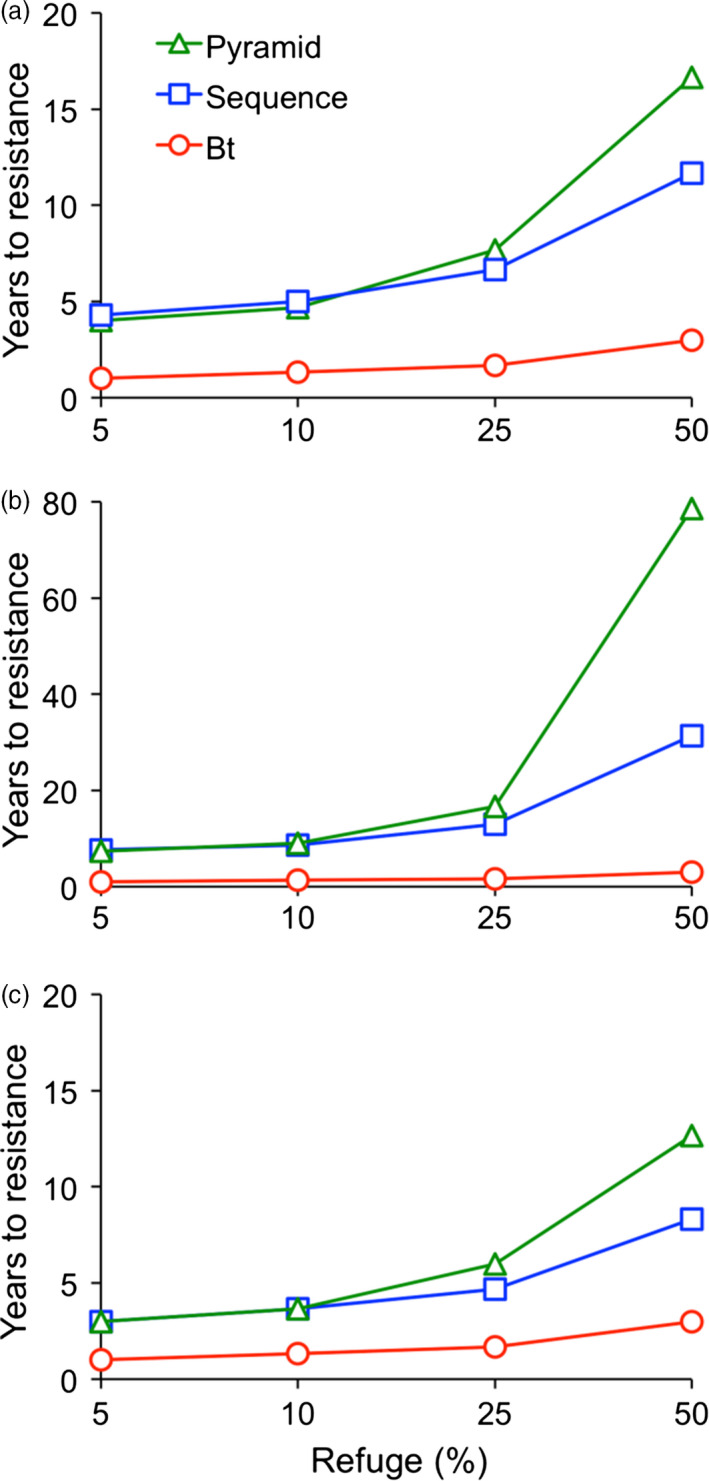
Transgenic crops producing insecticidal proteins from the bacterium Bacillus thuringiensis (Bt) are extensively cultivated worldwide. To counter rapidly increasing pest resistance to crops that produce single Bt toxins, transgenic plant 'pyramids' producing two or more Bt toxins that kill the same pest have been widely adopted. However, cross-resistance and antagonism between Bt toxins limit the sustainability of this approach. Here we describe development and testing of the first pyramids of cotton combining protection from a Bt toxin and RNA interference (RNAi). We developed two types of transgenic cotton plants producing double-stranded RNA (dsRNA) from the global lepidopteran pest Helicoverpa armigera designed to interfere with its metabolism of juvenile hormone (JH). We focused on suppression of JH acid methyltransferase (JHAMT), which is crucial for JH synthesis, and JH-binding protein (JHBP), which transports JH to organs. In 2015 and 2016, we tested larvae from a Bt-resistant strain and a related susceptible strain of H. armigera on seven types of cotton: two controls, Bt cotton, two types of RNAi cotton (targeting JHAMT or JHBP) and two pyramids (Bt cotton plus each type of RNAi). Both types of RNAi cotton were effective against Bt-resistant insects. Bt cotton and RNAi acted independently against the susceptible strain. In computer simulations of conditions in northern China, where millions of farmers grow Bt cotton as well as abundant non-transgenic host plants of H. armigera, pyramided cotton combining a Bt toxin and RNAi substantially delayed resistance relative to using Bt cotton alone.
Keywords: Bacillus thuringiensis; Helicoverpa armigera; RNA interference; genetic engineering; juvenile hormone; sustainability.
© 2017 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Conflict of interest statement
Z.Z. and Xiaofang Wang have a Chinese patent application related to this work, number CN102464710A, ‘
Figures
References
-
- Alstad, D. (2005) Populus Java Version 5.4. Department of Ecology, Evolution, and Behavior, University of Minnesota. Available at: http://www.cbs.umn.edu/populus [Accessed 2 September, 2016].
-
- Asokan, R. , Sharath Chandra, G. , Manamohan, M. , Krishna Kumar, N.K. and Sita, T. (2014) Response of various target genes to diet‐delivered dsRNA mediated RNA interference in the cotton bollworm, Helicoverpa armigera . J. Pest Sci. 87, 163–172.
-
- Baum, J.A. and Roberts, J.K. (2014) Progress towards RNAi‐mediated pest management. Adv. Insect Physiol. 47, 249–295.
-
- Baum, J.A. , Bogaert, T. , Clinton, W. , Heck, G.R. , Feldmann, P. , Ilagan, O. , Johnson, S. et al. (2007) Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 25, 1322–1326. - PubMed
-
- Belles, X. , Martin, D. and Piulachs, M.D. (2005) The mevalonate pathway and the synthesis of juvenile hormone in insects. Annu. Rev. Entomol. 50, 181–199. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
