

The SIN3A/HDAC Corepressor Complex Functionally Cooperates with NANOG to Promote Pluripotency
- PMID: 28199843
- PMCID: PMC5328122
- DOI: 10.1016/j.celrep.2017.01.055
The SIN3A/HDAC Corepressor Complex Functionally Cooperates with NANOG to Promote Pluripotency
Abstract
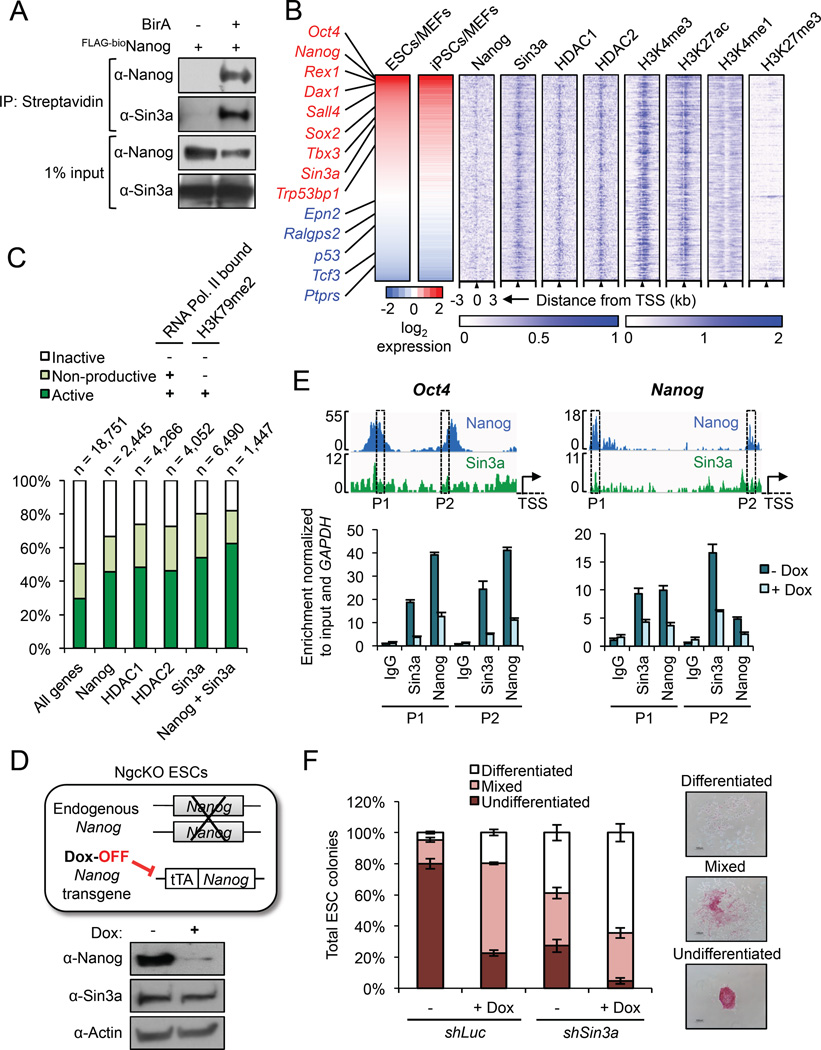
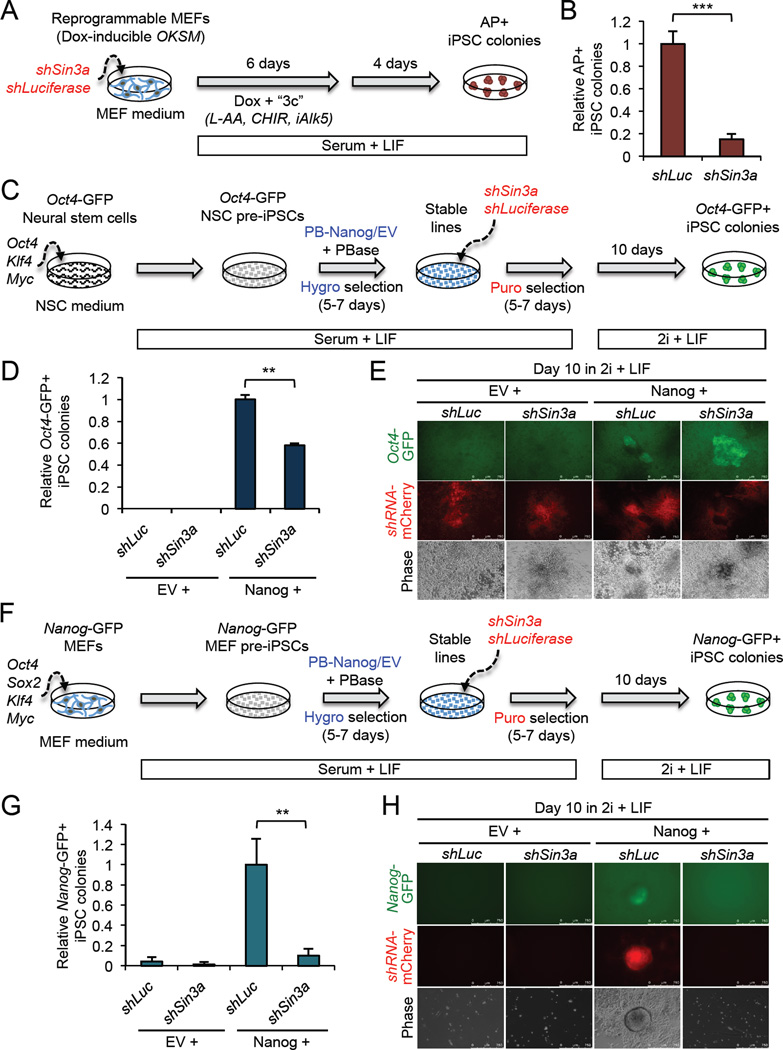
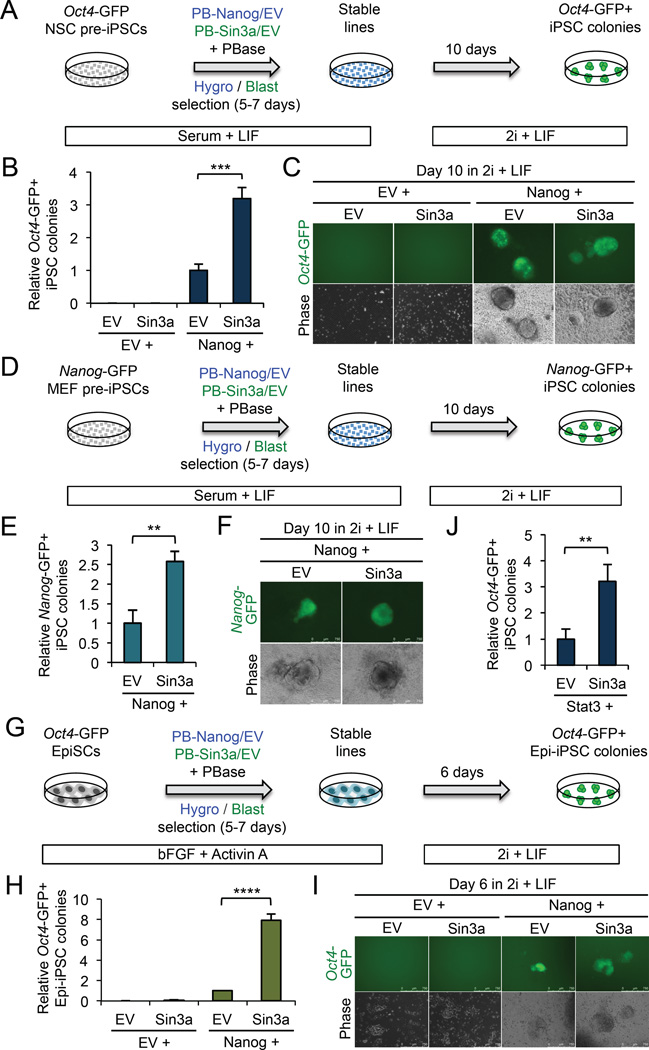
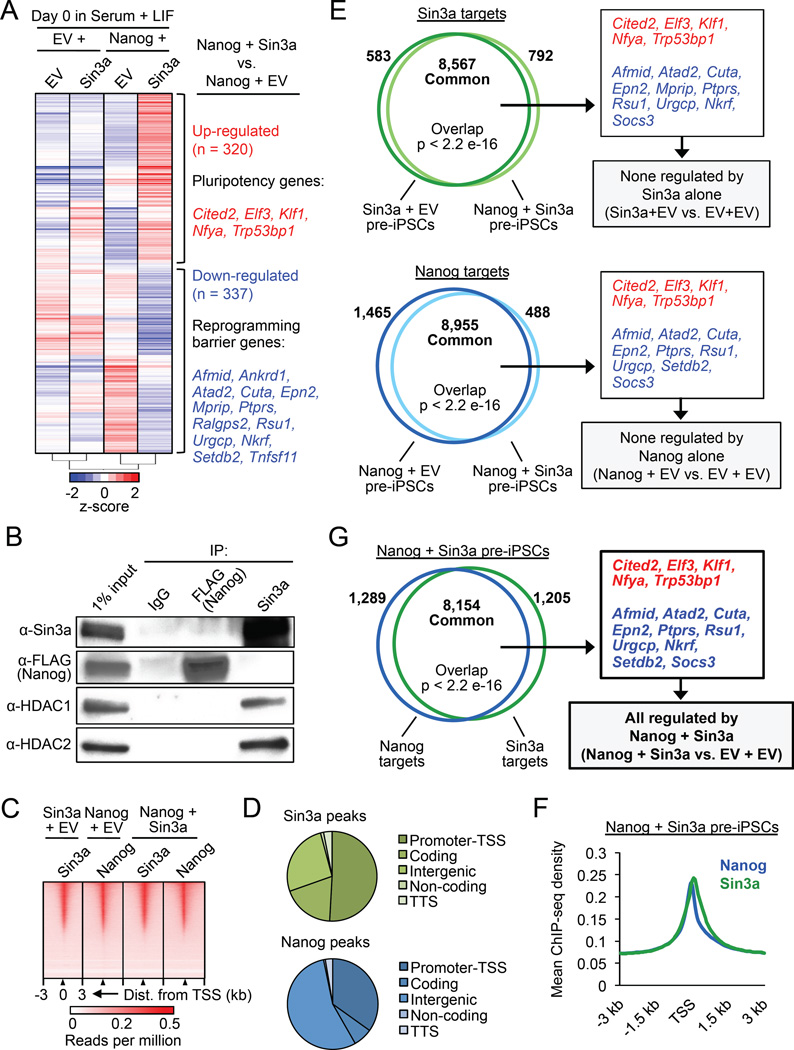
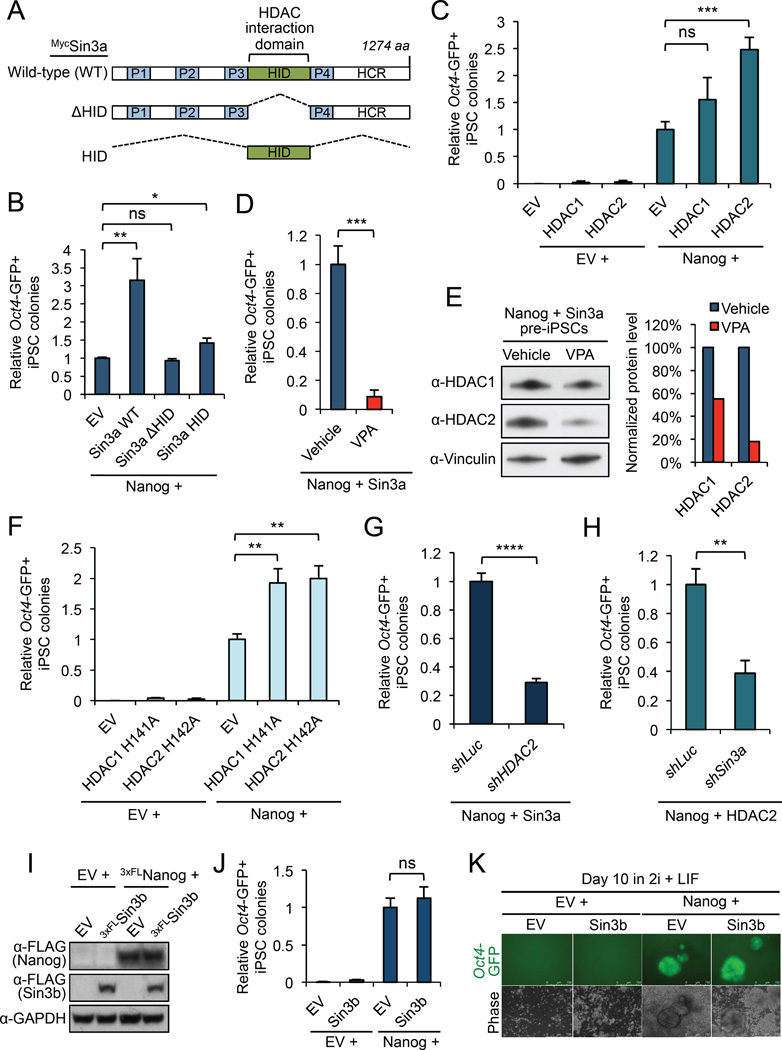
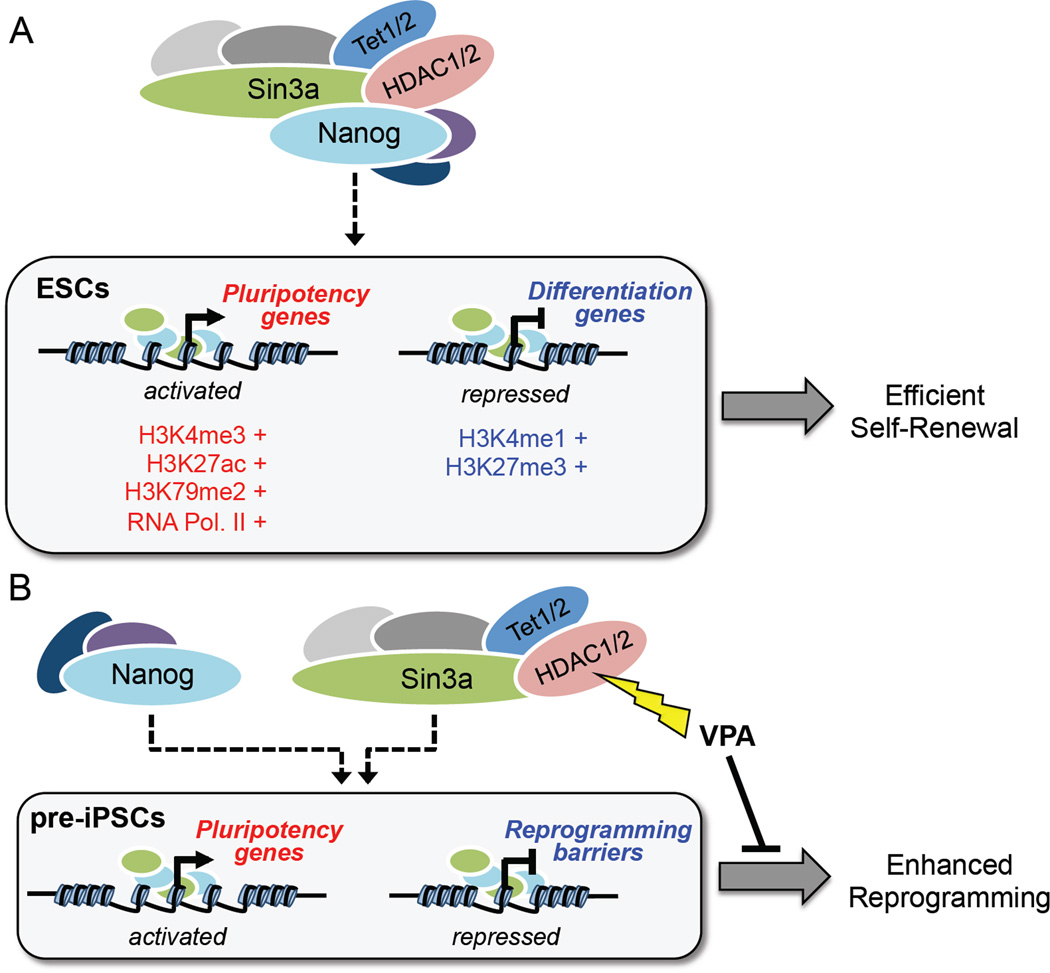
Although SIN3A is required for the survival of early embryos and embryonic stem cells (ESCs), the role of SIN3A in the maintenance and establishment of pluripotency remains unclear. Here, we find that the SIN3A/HDAC corepressor complex maintains ESC pluripotency and promotes the generation of induced pluripotent stem cells (iPSCs). Members of the SIN3A/HDAC corepressor complex are enriched in an extended NANOG interactome and function in transcriptional coactivation in ESCs. We also identified a critical role for SIN3A and HDAC2 in efficient reprogramming of somatic cells. Mechanistically, NANOG and SIN3A co-occupy transcriptionally active pluripotency genes in ESCs and also co-localize extensively at their genome-wide targets in pre-iPSCs. Additionally, both factors are required to directly induce a synergistic transcriptional program wherein pluripotency genes are activated and reprogramming barrier genes are repressed. Our findings indicate a transcriptional regulatory role for a major HDAC-containing complex in promoting pluripotency.
Keywords: ESCs; EpiSCs; HDAC1; HDAC2; SIN3A; SIN3B; TET1/2; VPA; iPSCs; pre-iPSCs.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
