

Genome-Wide Identification of Basic Helix-Loop-Helix and NF-1 Motifs Underlying GR Binding Sites in Male Rat Hippocampus
- PMID: 28200020
- PMCID: PMC5460825
- DOI: 10.1210/en.2016-1929
Genome-Wide Identification of Basic Helix-Loop-Helix and NF-1 Motifs Underlying GR Binding Sites in Male Rat Hippocampus
Abstract
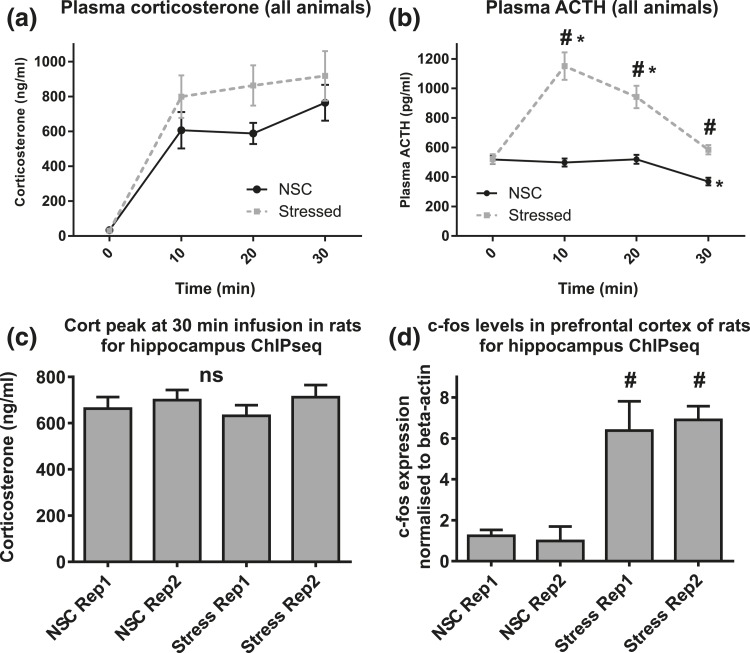
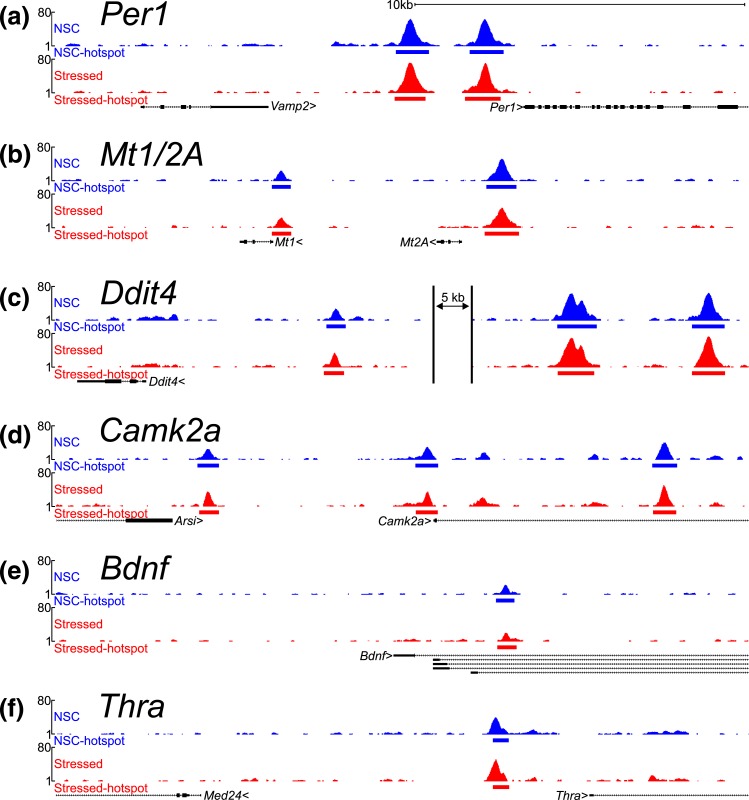
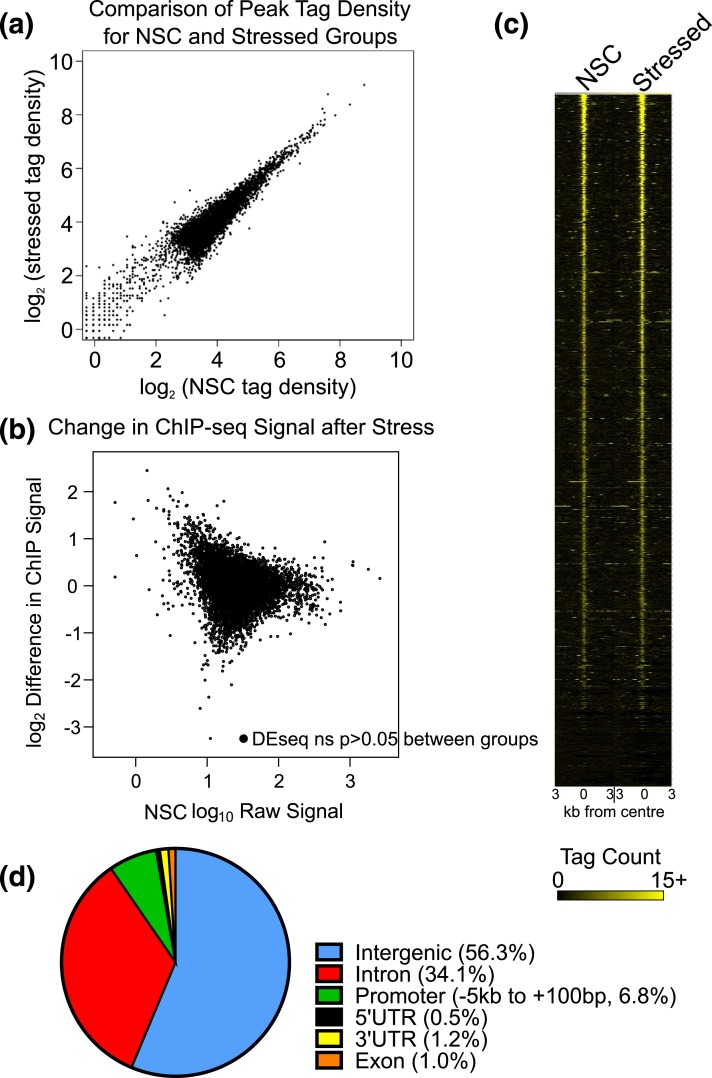
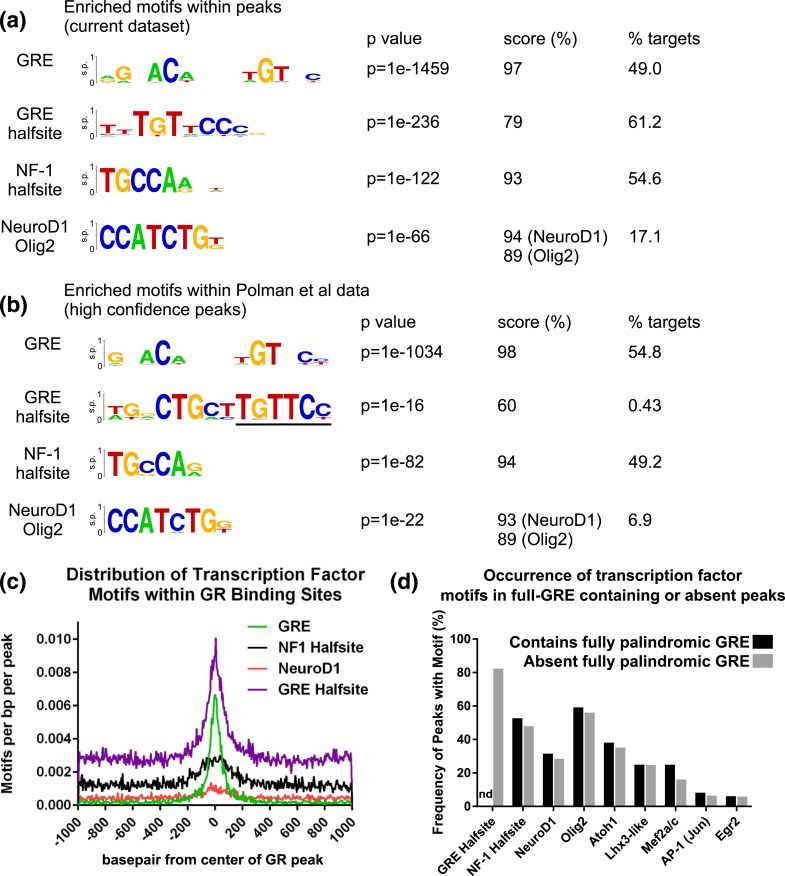
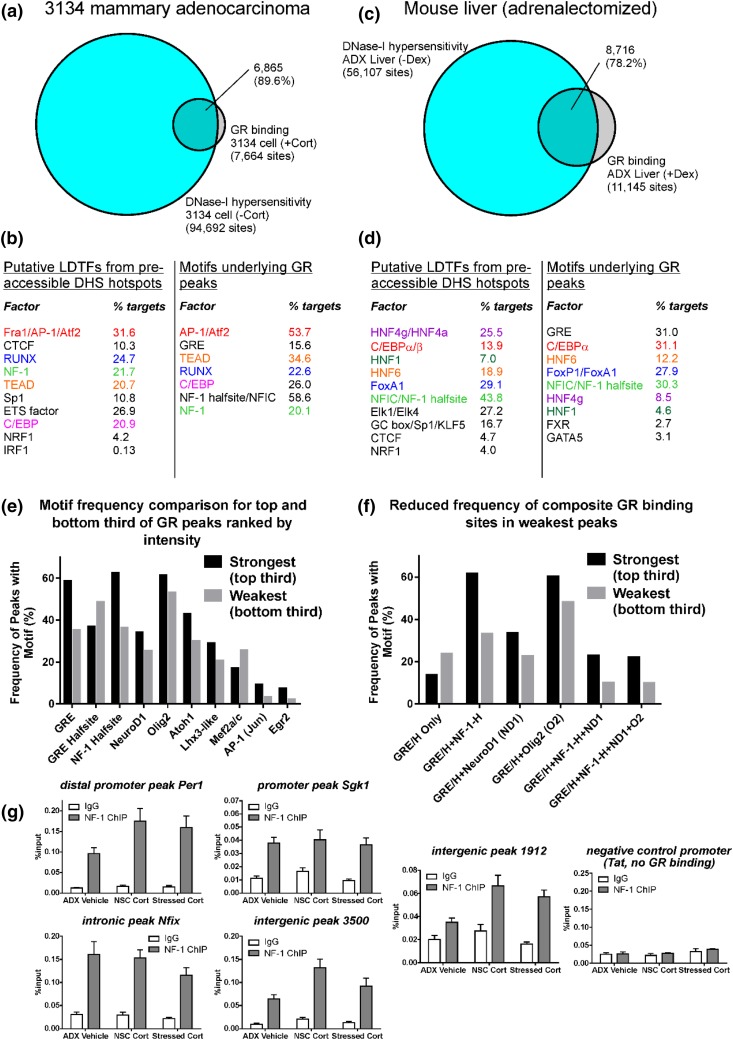
Glucocorticoids regulate hippocampal function in part by modulating gene expression through the glucocorticoid receptor (GR). GR binding is highly cell type specific, directed to accessible chromatin regions established during tissue differentiation. Distinct classes of GR binding sites are dependent on the activity of additional signal-activated transcription factors that prime chromatin toward context-specific organization. We hypothesized a stress context dependency for GR binding in hippocampus as a consequence of rapidly induced stress mediators priming chromatin accessibility. Using chromatin immunoprecipitation sequencing to interrogate GR binding, we found no effect of restraint stress context on GR binding, although analysis of sequences underlying GR binding sites revealed mechanistic detail for hippocampal GR function. We note enrichment of GR binding sites proximal to genes linked to structural and organizational roles, an absence of major tethering partners for GRs, and little or no evidence for binding at negative glucocorticoid response elements. A basic helix-loop-helix motif closely resembling a NeuroD1 or Olig2 binding site was found underlying a subset of GR binding sites and is proposed as a candidate lineage-determining transcription factor directing hippocampal chromatin access for GRs. Of our GR binding sites, 54% additionally contained half-sites for nuclear factor (NF)-1 that we propose as a collaborative or general transcription factor involved in hippocampal GR function. Our findings imply a dose-dependent and context-independent action of GRs in the hippocampus. Alterations in the expression or activity of NF-1/basic helix-loop-helix factors may play an as yet undetermined role in glucocorticoid-related disease susceptibility and outcome by altering GR access to hippocampal binding sites.
Figures
Similar articles
-
NeuroD Factors Discriminate Mineralocorticoid From Glucocorticoid Receptor DNA Binding in the Male Rat Brain.Endocrinology. 2017 May 1;158(5):1511-1522. doi: 10.1210/en.2016-1422. Endocrinology. 2017. PMID: 28324065
-
A genome-wide signature of glucocorticoid receptor binding in neuronal PC12 cells.BMC Neurosci. 2012 Oct 3;13:118. doi: 10.1186/1471-2202-13-118. BMC Neurosci. 2012. PMID: 23031785 Free PMC article.
-
Specific regulatory motifs predict glucocorticoid responsiveness of hippocampal gene expression.Endocrinology. 2011 Oct;152(10):3749-57. doi: 10.1210/en.2011-0287. Epub 2011 Aug 16. Endocrinology. 2011. PMID: 21846803
-
How glucocorticoid receptors modulate the activity of other transcription factors: a scope beyond tethering.Mol Cell Endocrinol. 2013 Nov 5;380(1-2):41-54. doi: 10.1016/j.mce.2012.12.014. Epub 2012 Dec 23. Mol Cell Endocrinol. 2013. PMID: 23267834 Review.
-
Genomic effects of glucocorticoids.Protoplasma. 2017 May;254(3):1175-1185. doi: 10.1007/s00709-016-1063-y. Epub 2016 Dec 24. Protoplasma. 2017. PMID: 28013411 Review.
Cited by
-
The evolving neurobiology of early-life stress.Neuron. 2025 May 21;113(10):1474-1490. doi: 10.1016/j.neuron.2025.02.016. Epub 2025 Mar 17. Neuron. 2025. PMID: 40101719 Review.
-
Experience and activity-dependent control of glucocorticoid receptors during the stress response in large-scale brain networks.Stress. 2021 Mar;24(2):130-153. doi: 10.1080/10253890.2020.1806226. Epub 2020 Aug 26. Stress. 2021. PMID: 32755268 Free PMC article.
-
Glucocorticoid and Mineralocorticoid Receptors in the Brain: A Transcriptional Perspective.J Endocr Soc. 2019 Jul 24;3(10):1917-1930. doi: 10.1210/js.2019-00158. eCollection 2019 Oct 1. J Endocr Soc. 2019. PMID: 31598572 Free PMC article. Review.
-
Coordinated transcriptional regulation by thyroid hormone and glucocorticoid interaction in adult mouse hippocampus-derived neuronal cells.PLoS One. 2019 Jul 26;14(7):e0220378. doi: 10.1371/journal.pone.0220378. eCollection 2019. PLoS One. 2019. PMID: 31348800 Free PMC article.
-
Do Corticosteroid Receptor mRNA Levels Predict the Expression of Their Target Genes?J Endocr Soc. 2022 Dec 12;7(2):bvac188. doi: 10.1210/jendso/bvac188. eCollection 2022 Dec 15. J Endocr Soc. 2022. PMID: 36578881 Free PMC article.
References
-
- McEwen BS. Physiology and neurobiology of stress and adaptation: central role of the brain. Physiol Rev. 2007;87(3):873–904. - PubMed
-
- Munck A, Guyre PM, Holbrook NJ. Physiological functions of glucocorticoids in stress and their relation to pharmacological actions. Endocr Rev. 1984;5(1):25–44. - PubMed
-
- Carroll, Curtis GC, Mendels J. Neuroendocrine regulation in depression. I. Limbic system-adrenocortical dysfunction. Arch Gen Psychiatry. 1976;33(9):1039–1044. - PubMed
-
- Yehuda R, Resnick H, Kahana B, Giller EL. Long-lasting hormonal alterations to extreme stress in humans: normative or maladaptive? Psychosom Med. 1993;55(3):287–297. - PubMed
-
- Condren RM, O’Neill A, Ryan MC, Barrett P, Thakore JH. HPA axis response to a psychological stressor in generalised social phobia. Psychoneuroendocrinology. 2002;27(6):693–703. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
