

Tiam1/Vav2-Rac1 axis: A tug-of-war between islet function and dysfunction
- PMID: 28202288
- PMCID: PMC5382093
- DOI: 10.1016/j.bcp.2017.02.007
Tiam1/Vav2-Rac1 axis: A tug-of-war between islet function and dysfunction
Abstract
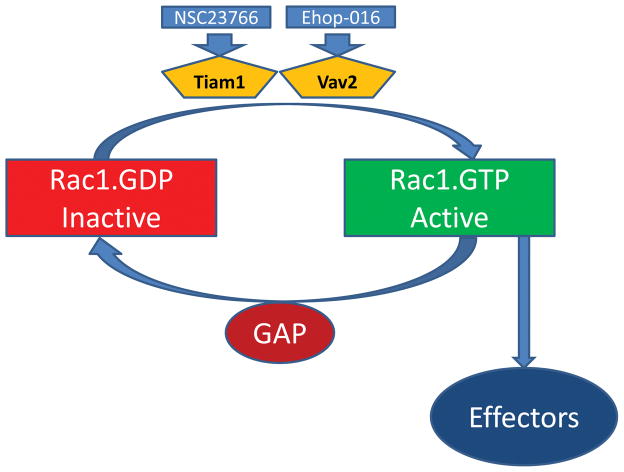
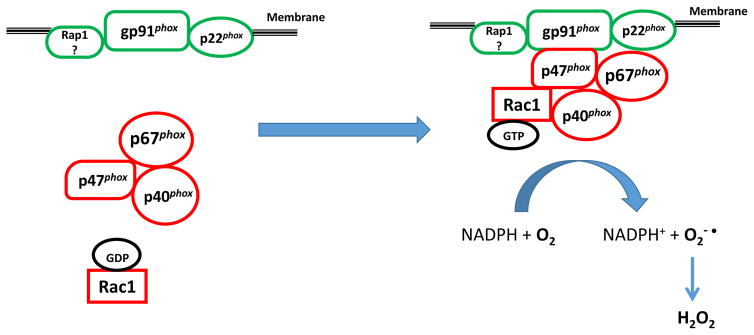
Glucose-stimulated insulin secretion [GSIS] from the islet β-cell involves a well-orchestrated interplay between metabolic and cationic events. It is well established that intracellular generation of adenine and guanine nucleotide triphosphates [e.g., ATP and GTP] represents one of the requisite signaling steps in GSIS. The small molecular mass GTP-binding proteins [G-proteins; e.g., Rac1 and Cdc42] have been shown to regulate islet β-cell function including actin cytoskeletal remodeling and fusion of insulin granules with the plasma membrane for GSIS to occur. In this context, several regulatory factors for these G-proteins have been identified in the pancreatic β-cell; these include guanine nucleotide exchange factors [GEFs] and guanine nucleotide dissociation inhibitors [GDI]. Recent pharmacological and molecular biological evidence identified Tiam1 and Vav2 as GEFs for Rac1 in promoting physiological insulin secretion. Paradoxically, emerging evidence in multiple cell types, including the islet β-cell, suggests key roles for Rac1 in the onset of cellular dysfunction under conditions of metabolic stress and diabetes. Furthermore, functional inactivation of either Tiam1 or Vav2 appears to attenuate sustained activation of Rac1 and its downstream signaling events [activation of stress kinases] under conditions of metabolic stress. Together, these findings suggest both "friendly" and "non-friendly" roles for Tiam1/Vav2-Rac1 signaling axis in islet β-cell in health and diabetes. Our current understanding of the field and the knowledge gaps that exist in this area of islet biology are heighted herein. Furthermore, potential caveats in the specificity and selectivity of pharmacological inhibitors that are available currently are discussed in this Commentary.
Keywords: G proteins; Glucotoxicity; Islet β-cell; Rac1; Tiam1; Vav2.
Published by Elsevier Inc.
Figures

Similar articles
-
Role of G-proteins in islet function in health and diabetes.Diabetes Obes Metab. 2017 Sep;19 Suppl 1(Suppl 1):63-75. doi: 10.1111/dom.13011. Diabetes Obes Metab. 2017. PMID: 28880478 Free PMC article. Review.
-
Friendly, and not so friendly, roles of Rac1 in islet β-cell function: lessons learnt from pharmacological and molecular biological approaches.Biochem Pharmacol. 2011 Apr 15;81(8):965-75. doi: 10.1016/j.bcp.2011.01.013. Epub 2011 Feb 12. Biochem Pharmacol. 2011. PMID: 21300027 Free PMC article.
-
VAV2, a guanine nucleotide exchange factor for Rac1, regulates glucose-stimulated insulin secretion in pancreatic beta cells.Diabetologia. 2015 Nov;58(11):2573-81. doi: 10.1007/s00125-015-3707-4. Epub 2015 Jul 31. Diabetologia. 2015. PMID: 26224100 Free PMC article.
-
Regulatory roles for Tiam1, a guanine nucleotide exchange factor for Rac1, in glucose-stimulated insulin secretion in pancreatic beta-cells.Biochem Pharmacol. 2009 Jan 1;77(1):101-13. doi: 10.1016/j.bcp.2008.09.021. Epub 2008 Sep 27. Biochem Pharmacol. 2009. PMID: 18930714 Free PMC article.
-
Roles of GTP and Rho GTPases in pancreatic islet beta cell function and dysfunction.Small GTPases. 2021 Sep-Nov;12(5-6):323-335. doi: 10.1080/21541248.2020.1815508. Epub 2020 Aug 31. Small GTPases. 2021. PMID: 32867592 Free PMC article. Review.
Cited by
-
Regulatory roles of CARD9-BCL10-Rac1 (CBR) signalome in islet β-cell function in health and metabolic stress: Is there room for MALT1?Biochem Pharmacol. 2023 Dec;218:115889. doi: 10.1016/j.bcp.2023.115889. Epub 2023 Oct 29. Biochem Pharmacol. 2023. PMID: 37991197 Free PMC article. Review.
-
Exploring the interaction between extracellular matrix components in a 3D organoid disease model to replicate the pathophysiology of breast cancer.J Exp Clin Cancer Res. 2023 Dec 16;42(1):343. doi: 10.1186/s13046-023-02926-4. J Exp Clin Cancer Res. 2023. PMID: 38102637 Free PMC article. Review.
-
Deimination Protein Profiles in Alligator mississippiensis Reveal Plasma and Extracellular Vesicle-Specific Signatures Relating to Immunity, Metabolic Function, and Gene Regulation.Front Immunol. 2020 Apr 28;11:651. doi: 10.3389/fimmu.2020.00651. eCollection 2020. Front Immunol. 2020. PMID: 32411128 Free PMC article.
-
Role of G-proteins in islet function in health and diabetes.Diabetes Obes Metab. 2017 Sep;19 Suppl 1(Suppl 1):63-75. doi: 10.1111/dom.13011. Diabetes Obes Metab. 2017. PMID: 28880478 Free PMC article. Review.
-
Tiam1 methylation by NSD2 promotes Rac1 signaling activation and colon cancer metastasis.Proc Natl Acad Sci U S A. 2023 Dec 26;120(52):e2305684120. doi: 10.1073/pnas.2305684120. Epub 2023 Dec 19. Proc Natl Acad Sci U S A. 2023. PMID: 38113258 Free PMC article.
References
-
- Prentki M, Matschinsky FM, Madiraju SR. Metabolic signaling in fuel-induced insulin secretion. Cell Metab. 2013;18:162–185. - PubMed
-
- Berggren PO, Leibiger IB. Novel aspects on signal transduction in the pancreatic beta cell. Nutr Metab Cardiovasc Dis. 2006;16(suppl 1):S7–S10. - PubMed
-
- Metz SA, Rabaglia ME, Pintar TJ. Selective inhibitors of GTP synthesis impede exocytotic insulin release from intact rat islets. J Biol Chem. 1992;267:12517–12527. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
