

A key centriole assembly interaction interface between human PLK4 and STIL appears to not be conserved in flies
- PMID: 28202467
- PMCID: PMC5374404
- DOI: 10.1242/bio.024661
A key centriole assembly interaction interface between human PLK4 and STIL appears to not be conserved in flies
Abstract
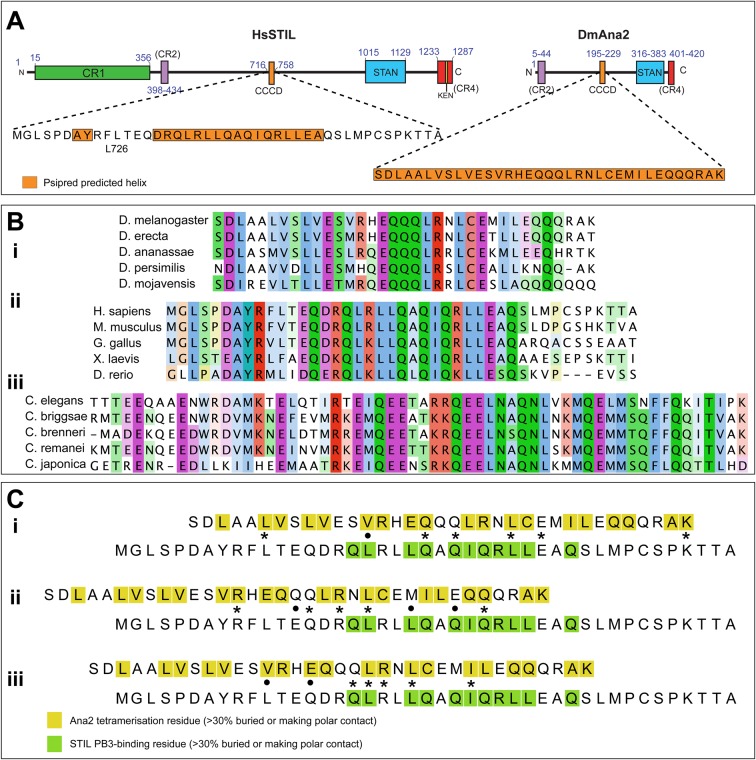
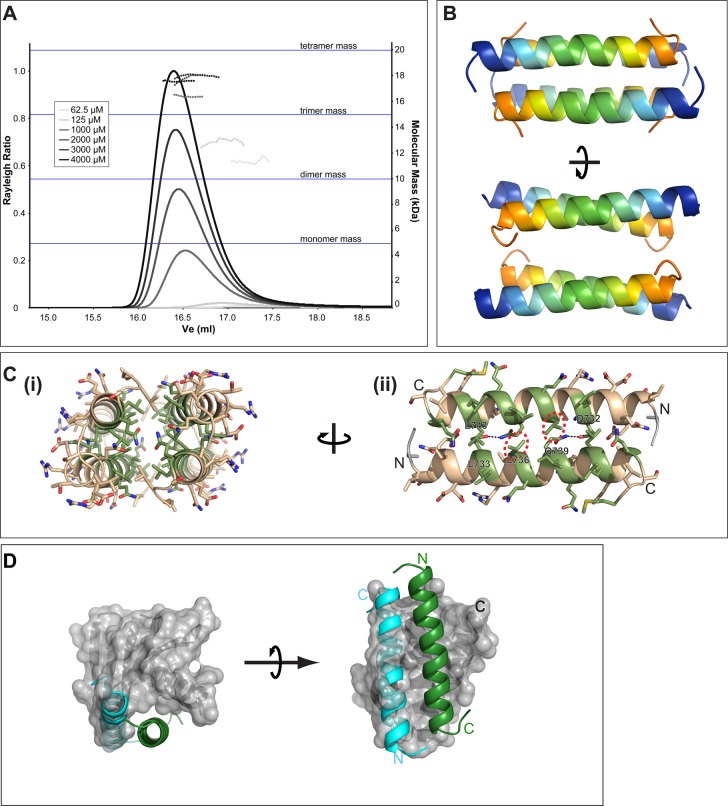
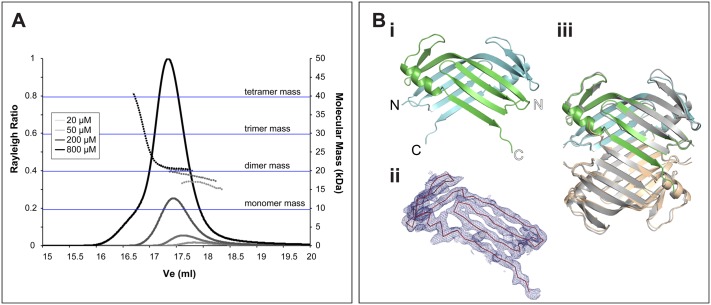
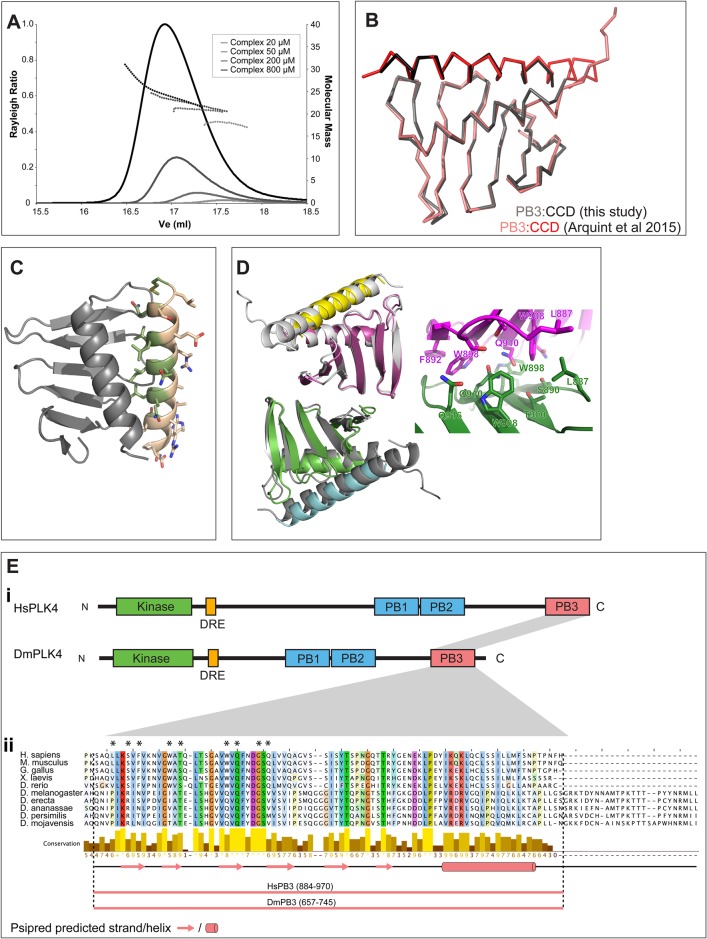
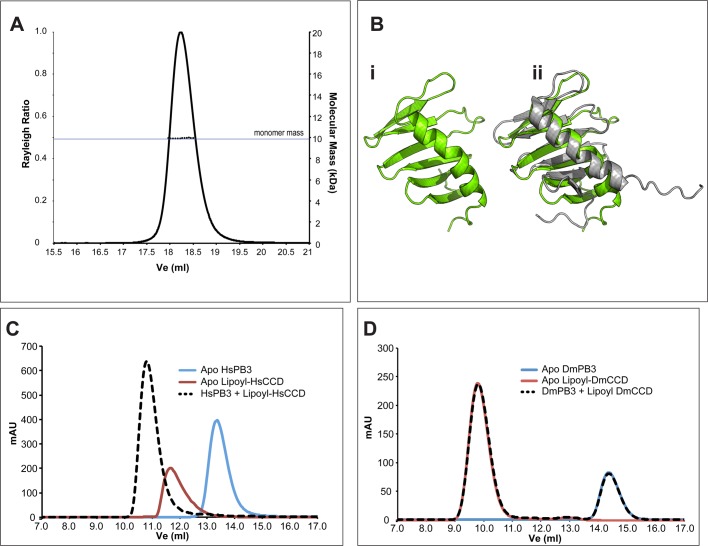
A small number of proteins form a conserved pathway of centriole duplication. In humans and flies, the binding of PLK4/Sak to STIL/Ana2 initiates daughter centriole assembly. In humans, this interaction is mediated by an interaction between the Polo-Box-3 (PB3) domain of PLK4 and the coiled-coil domain of STIL (HsCCD). We showed previously that the Drosophila Ana2 coiled-coil domain (DmCCD) is essential for centriole assembly, but it forms a tight parallel tetramer in vitro that likely precludes an interaction with PB3. Here, we show that the isolated HsCCD and HsPB3 domains form a mixture of homo-multimers in vitro, but these readily dissociate when mixed to form the previously described 1:1 HsCCD:HsPB3 complex. In contrast, although Drosophila PB3 (DmPB3) adopts a canonical polo-box fold, it does not detectably interact with DmCCD in vitro Thus, surprisingly, a key centriole assembly interaction interface appears to differ between humans and flies.
Keywords: Cartwheel; Centriole duplication; Centrosome.
© 2017. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
Similar articles
-
An ordered pattern of Ana2 phosphorylation by Plk4 is required for centriole assembly.J Cell Biol. 2018 Apr 2;217(4):1217-1231. doi: 10.1083/jcb.201605106. Epub 2018 Mar 1. J Cell Biol. 2018. PMID: 29496738 Free PMC article.
-
The homo-oligomerisation of both Sas-6 and Ana2 is required for efficient centriole assembly in flies.Elife. 2015 May 23;4:e07236. doi: 10.7554/eLife.07236. Elife. 2015. PMID: 26002084 Free PMC article.
-
Plk4-dependent phosphorylation of STIL is required for centriole duplication.Biol Open. 2015 Feb 20;4(3):370-7. doi: 10.1242/bio.201411023. Biol Open. 2015. PMID: 25701666 Free PMC article.
-
The centrosome duplication cycle in health and disease.FEBS Lett. 2014 Aug 1;588(15):2366-72. doi: 10.1016/j.febslet.2014.06.030. Epub 2014 Jun 18. FEBS Lett. 2014. PMID: 24951839 Review.
-
Centrosome duplication: of rules and licenses.Trends Cell Biol. 2007 May;17(5):215-21. doi: 10.1016/j.tcb.2007.03.003. Epub 2007 Mar 26. Trends Cell Biol. 2007. PMID: 17383880 Review.
Cited by
-
An ordered pattern of Ana2 phosphorylation by Plk4 is required for centriole assembly.J Cell Biol. 2018 Apr 2;217(4):1217-1231. doi: 10.1083/jcb.201605106. Epub 2018 Mar 1. J Cell Biol. 2018. PMID: 29496738 Free PMC article.
-
A Role for Polo-Like Kinase 4 in Vascular Fibroblast Cell-Type Transition.JACC Basic Transl Sci. 2021 Mar 22;6(3):257-283. doi: 10.1016/j.jacbts.2020.12.015. eCollection 2021 Mar. JACC Basic Transl Sci. 2021. PMID: 33778212 Free PMC article.
-
Direct binding of CEP85 to STIL ensures robust PLK4 activation and efficient centriole assembly.Nat Commun. 2018 Apr 30;9(1):1731. doi: 10.1038/s41467-018-04122-x. Nat Commun. 2018. PMID: 29712910 Free PMC article.
-
The kinase ZYG-1 phosphorylates the cartwheel protein SAS-5 to drive centriole assembly in C. elegans.EMBO Rep. 2024 Jun;25(6):2698-2721. doi: 10.1038/s44319-024-00157-y. Epub 2024 May 14. EMBO Rep. 2024. PMID: 38744971 Free PMC article.
-
Interaction between the Caenorhabditis elegans centriolar protein SAS-5 and microtubules facilitates organelle assembly.Mol Biol Cell. 2018 Mar 15;29(6):722-735. doi: 10.1091/mbc.E17-06-0412. Epub 2018 Jan 24. Mol Biol Cell. 2018. PMID: 29367435 Free PMC article.
References
-
- Afonine P. V., Grosse-Kunstleve R. W., Echols N., Headd J. J., Moriarty N. W., Mustyakimov M., Terwilliger T. C., Urzhumtsev A., Zwart P. H. and Adams P. D. (2012). Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. D Biol. Crystallogr. 68, 352-367. 10.1107/S0907444912001308 - DOI - PMC - PubMed
-
- Chen V. B., Arendall W. B., Headd J. J., Keedy D. A., Immormino R. M., Kapral G. J., Murray L. W., Richardson J. S. and Richardson D. C. (2010). MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 66, 12-21. 10.1107/S0907444909042073 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
