

Chronic hypoxia upregulates DNA methyltransferase and represses large conductance Ca2+-activated K+ channel function in ovine uterine arteries
- PMID: 28203702
- PMCID: PMC6366546
- DOI: 10.1095/biolreprod.116.145946
Chronic hypoxia upregulates DNA methyltransferase and represses large conductance Ca2+-activated K+ channel function in ovine uterine arteries
Abstract
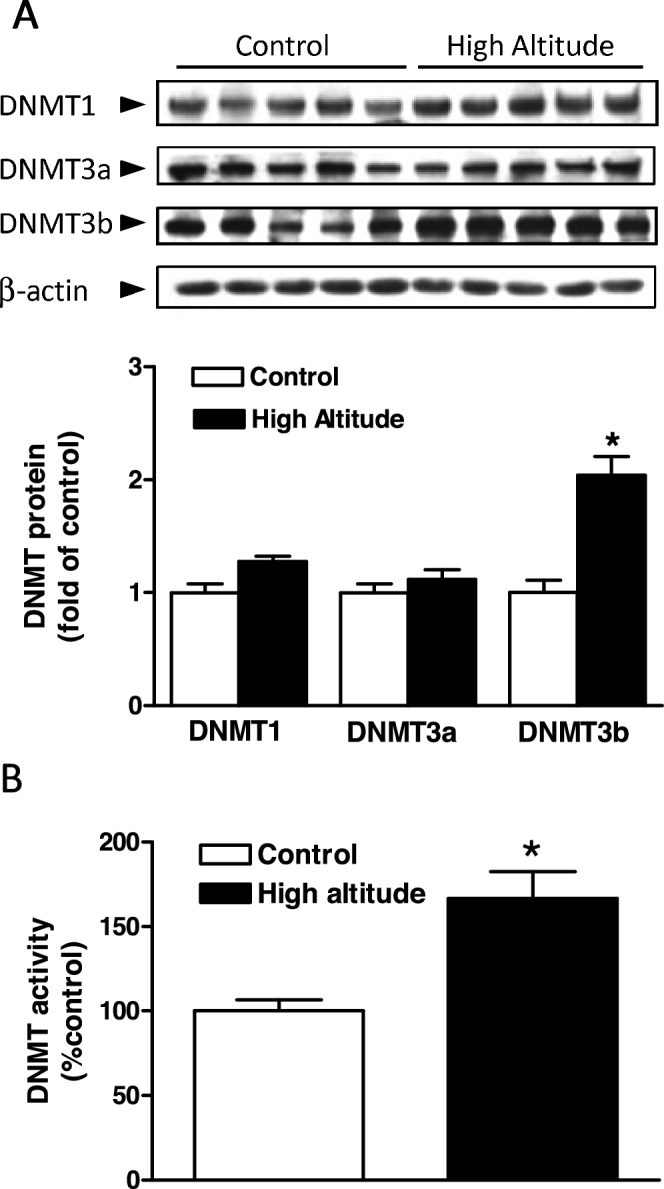
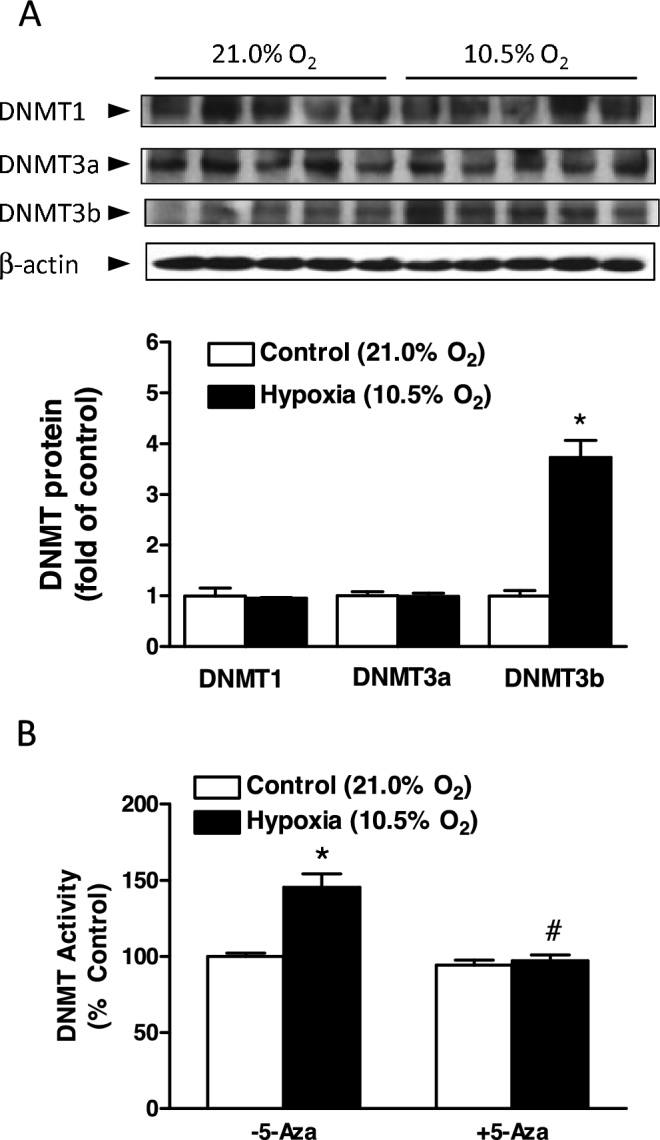
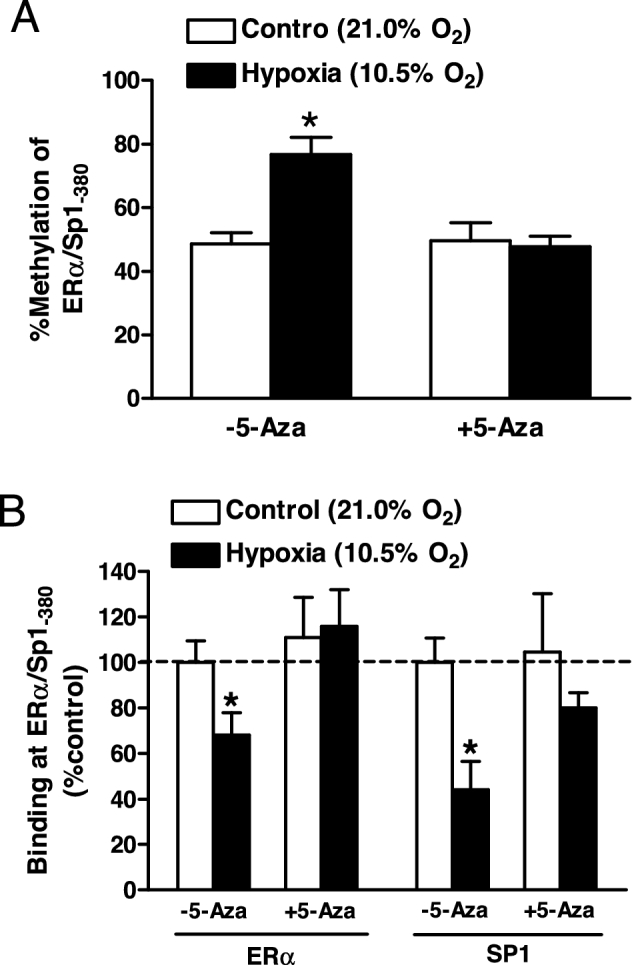
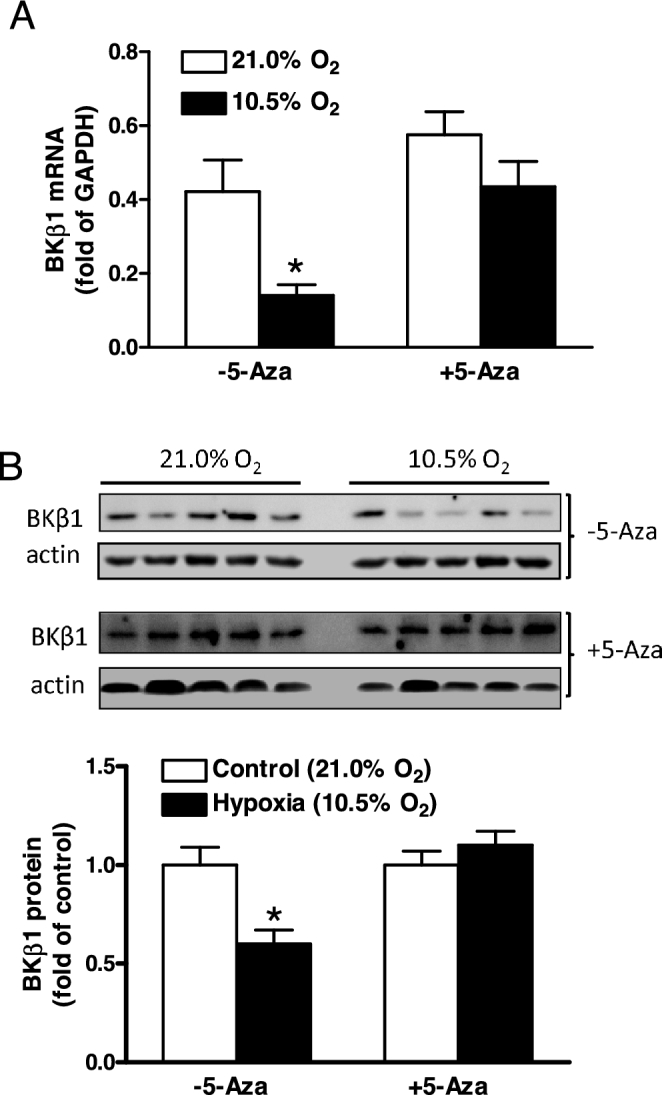
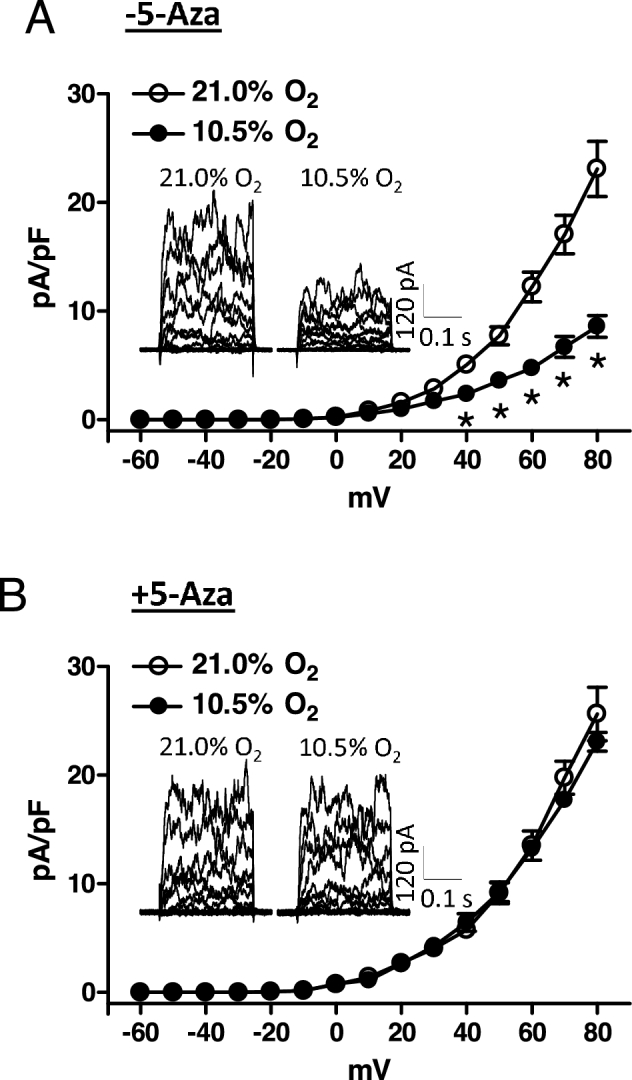
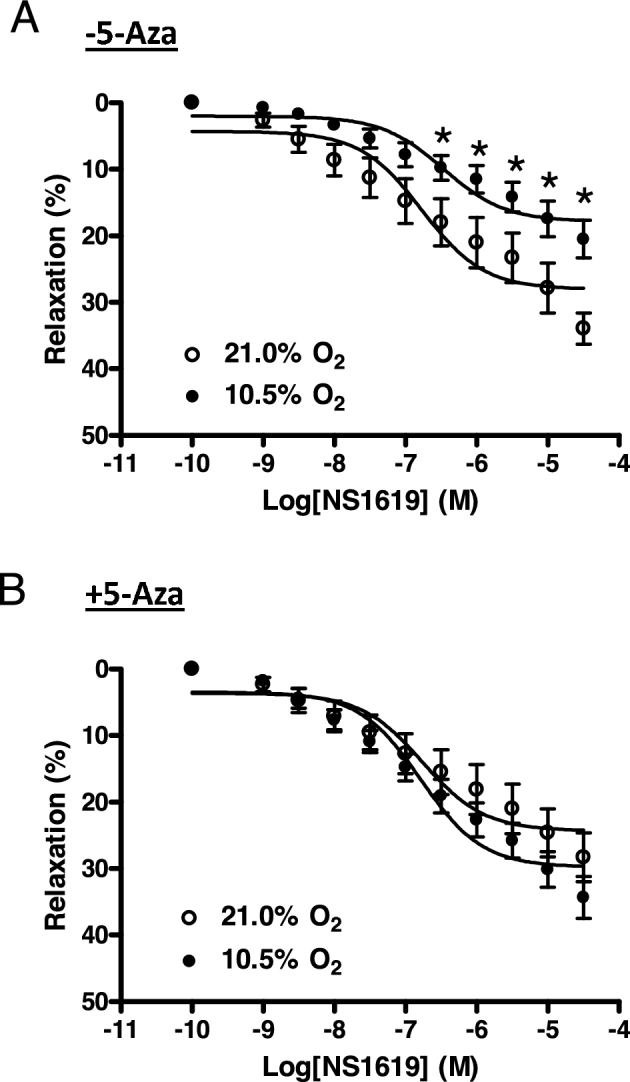
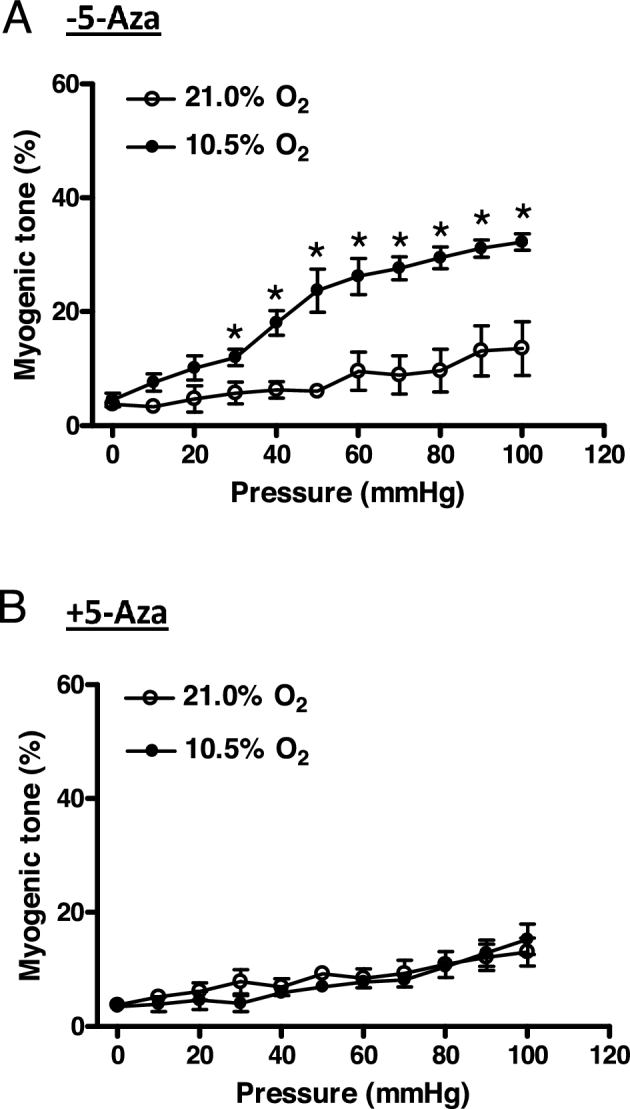
Chronic hypoxia during gestation suppresses large-conductance Ca2+-activated K+ (BKCa) channel function and impedes uterine arterial adaptation to pregnancy. This study tested the hypothesis that chronic hypoxia has a direct effect in upregulating DNA methyltransferase (DNMT) and epigenetically repressing BKCa channel beta-1 subunit (KCNMB1) expression in uterine arteries. Resistance-sized uterine arteries were isolated from near-term pregnant sheep maintained at ∼300 m above sea level or animals acclimatized to high-altitude (3,801 m) hypoxia for 110 days during gestation. For ex vivo hypoxia treatment, uterine arteries from normoxic animals were treated with 21.0% O2 or 10.5% O2 for 48 h. High-altitude hypoxia significantly upregulated DNMT3b expression and enzyme activity in uterine arteries. Similarly, ex vivo hypoxia treatment upregulated DNMT3b expression and enzyme activity that was blocked by a DNMT inhibitor 5-aza-2'-deoxycytidine (5-Aza). Of importance, 5-Aza inhibited hypoxia-induced hypermethylation of specificity protein (SP) 1 binding site at the KCNMB1 promoter and restored transcription factor binding to the KCNMB1 promoter, resulting in the recovery of KCNMB1 gene expression in uterine arteries. Furthermore, 5-Aza blocked the effect of hypoxia and rescued BKCa channel activity and reversed hypoxia-induced decrease in BKCa channel-mediated relaxations and increase in myogenic tone of uterine arteries. Collectively, these results suggest that chronic hypoxia during gestation upregulates DNMT expression and activity, resulting in hypermethylation and repression of KCNMB1 gene and BKCa channel function, impeding uterine arterial adaptation to pregnancy.
Keywords: hypoxia; DNA methylation; BKCa channel; myogenic tone; uterine arteries.
Figures
References
-
- Rosenfeld CR. Distribution of cardiac output in ovine pregnancy. Am J Physiol, 1977, 232:H231–H235. - PubMed
-
- Palmer SK, Zamudio S, Coffin C, Parker S, Stamm E, Moore LG. Quantitative estimation of human uterine artery blood flow and pelvic blood flow redistribution in pregnancy. Obstet Gynecol, 1992, 80:1000–1006. - PubMed
-
- Lang U, Baker RS, Braems G, Zygmunt M, Kunzel W, Clark KE. Uterine blood flow—a determinant of fetal growth. Eur J Obstet Gynecol Reprod Biol, 2003, 110, (Suppl 1):S5–S61. - PubMed
-
- Lambert G, Brichant JF, Hartstein G, Bonhomme V, Dewandre PY. Preeclampsia: an update. Acta Anaesthesiol Belg, 2014, 65:137–149. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
