

Tailored protein encapsulation into a DNA host using geometrically organized supramolecular interactions
- PMID: 28205515
- PMCID: PMC5316895
- DOI: 10.1038/ncomms14472
Tailored protein encapsulation into a DNA host using geometrically organized supramolecular interactions
Abstract
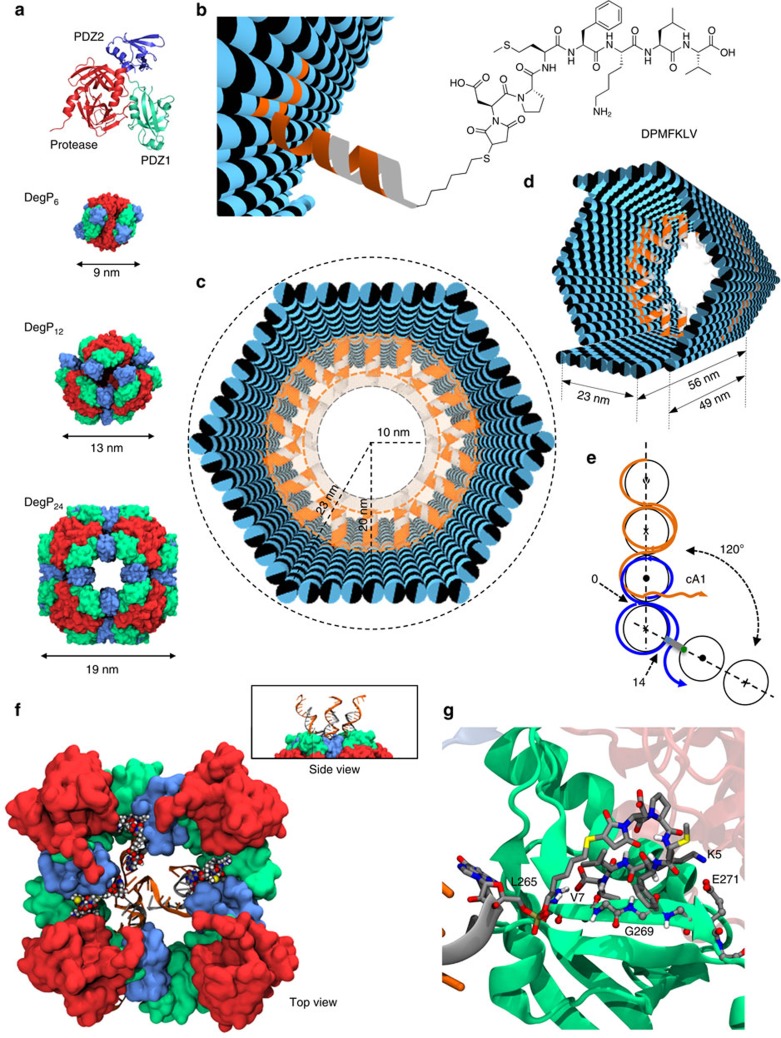
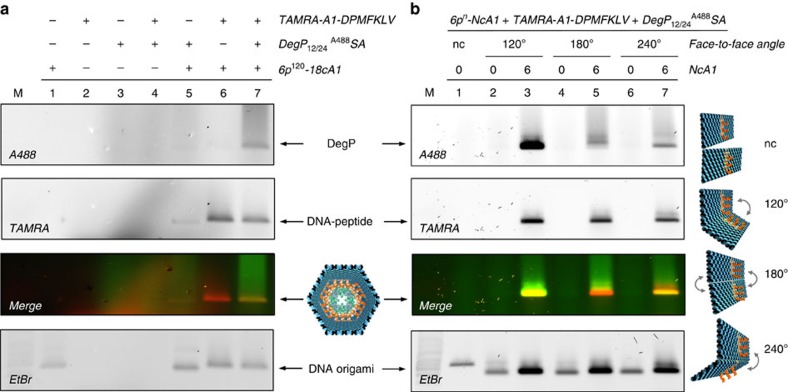
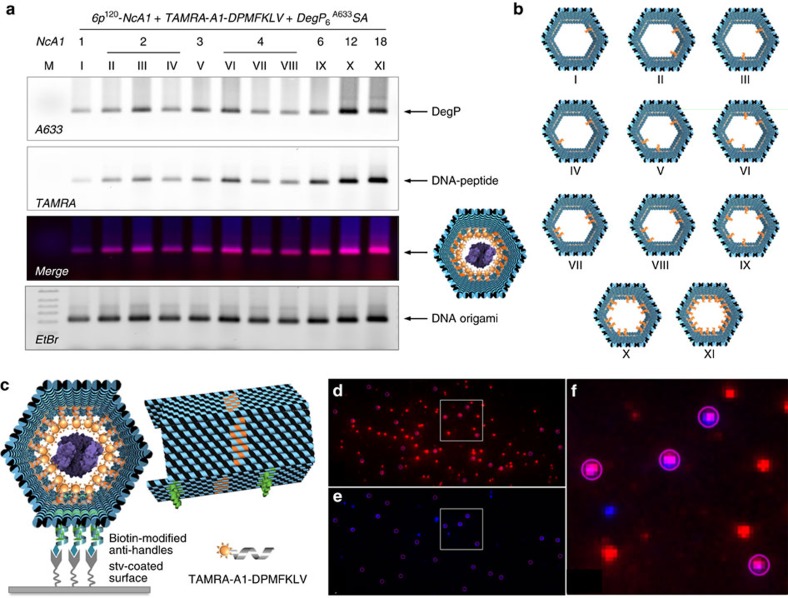
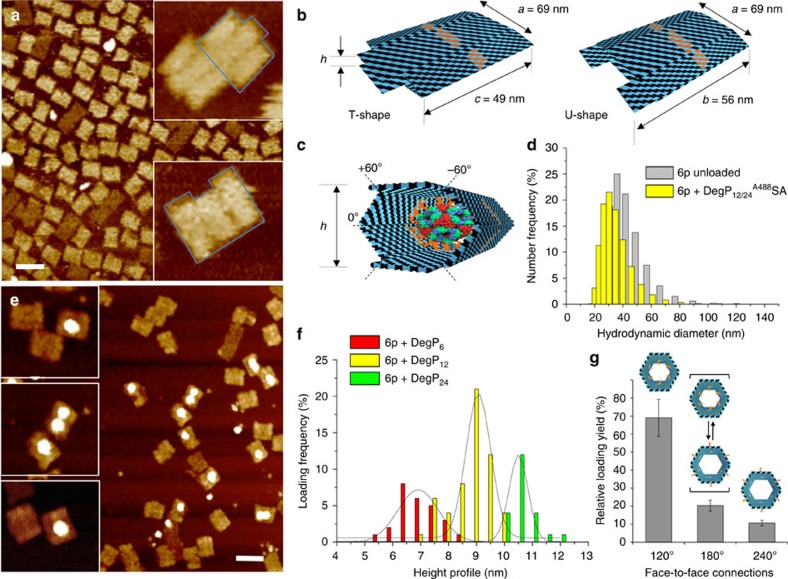
The self-organizational properties of DNA have been used to realize synthetic hosts for protein encapsulation. However, current strategies of DNA-protein conjugation still limit true emulation of natural host-guest systems, whose formation relies on non-covalent bonds between geometrically matching interfaces. Here we report one of the largest DNA-protein complexes of semisynthetic origin held in place exclusively by spatially defined supramolecular interactions. Our approach is based on the decoration of the inner surface of a DNA origami hollow structure with multiple ligands converging to their corresponding binding sites on the protein surface with programmable symmetry and range-of-action. Our results demonstrate specific host-guest recognition in a 1:1 stoichiometry and selectivity for the guest whose size guarantees sufficient molecular diffusion preserving short intermolecular distances. DNA nanocontainers can be thus rationally designed to trap single guest molecules in their native form, mimicking natural strategies of molecular recognition and anticipating a new method of protein caging.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

References
-
- Hurtley S. Spatial cell biology. Location, location, location. Introduction. Science 326, 1205 (2009). - PubMed
-
- Agapakis C. M., Boyle P. M. & Silver P. A. Natural strategies for the spatial optimization of metabolism in synthetic biology. Nat. Chem. Biol. 8, 527–535 (2012). - PubMed
-
- Chen A. H. & Silver P. A. Designing biological compartmentalization. Trends Cell Biol. 22, 662–670 (2012). - PubMed
-
- Conrado R. J., Varner J. D. & DeLisa M. P. Engineering the spatial organization of metabolic enzymes: mimicking nature's synergy. Curr. Opin. Biotechnol. 19, 492–499 (2008). - PubMed
-
- Lee H., DeLoache W. C. & Dueber J. E. Spatial organization of enzymes for metabolic engineering. Metab. Eng. 14, 242–251 (2012). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
