

Cationic amino acid transporters play key roles in the survival and transmission of apicomplexan parasites
- PMID: 28205520
- PMCID: PMC5316894
- DOI: 10.1038/ncomms14455
Cationic amino acid transporters play key roles in the survival and transmission of apicomplexan parasites
Abstract
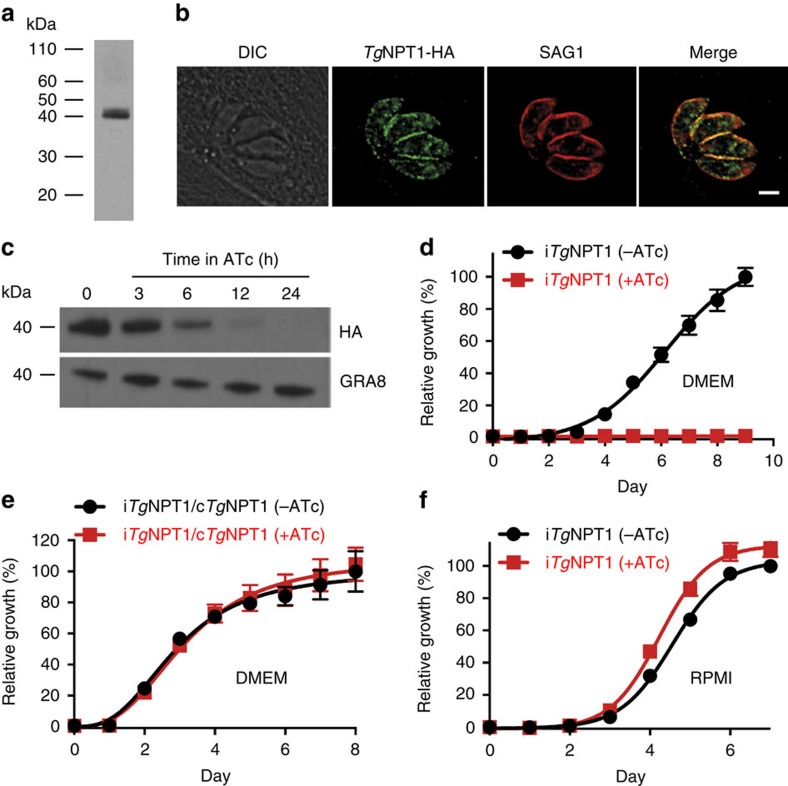
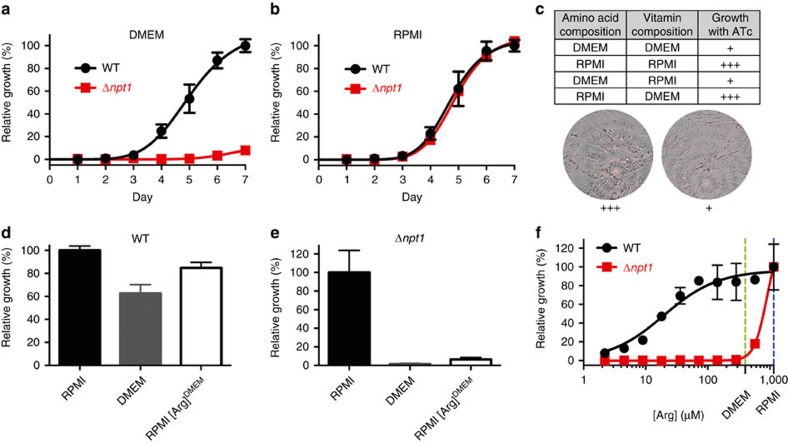
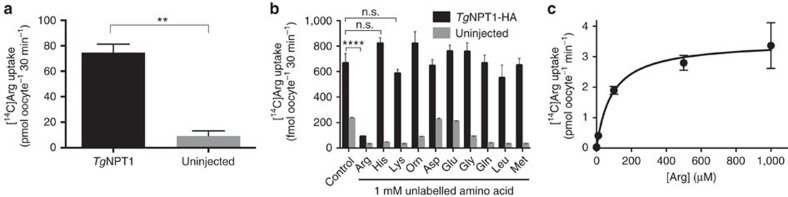
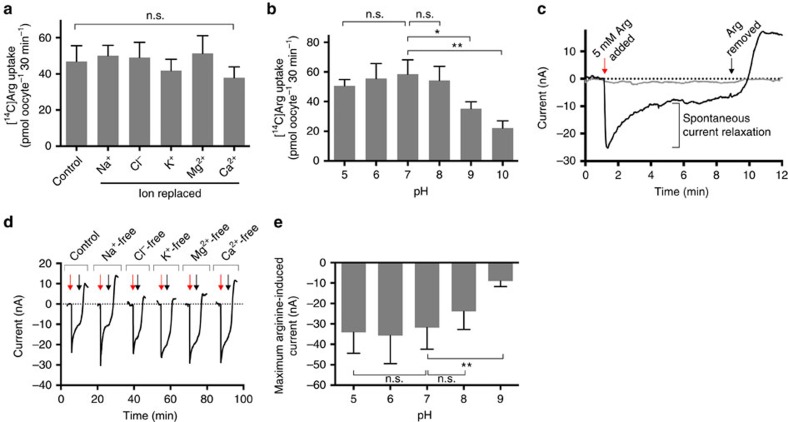
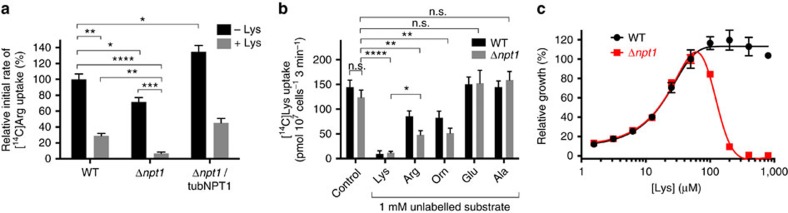
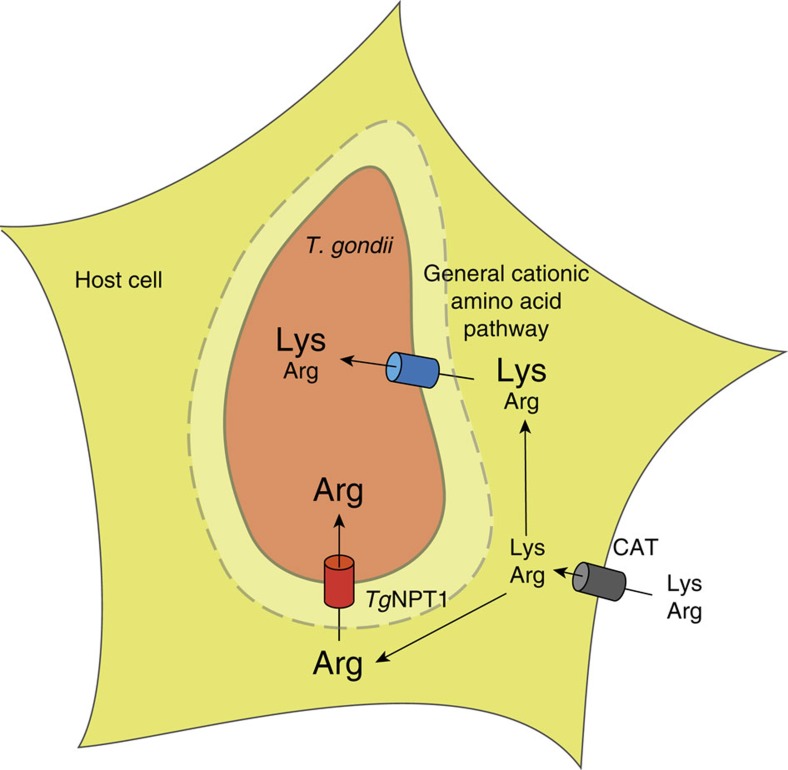
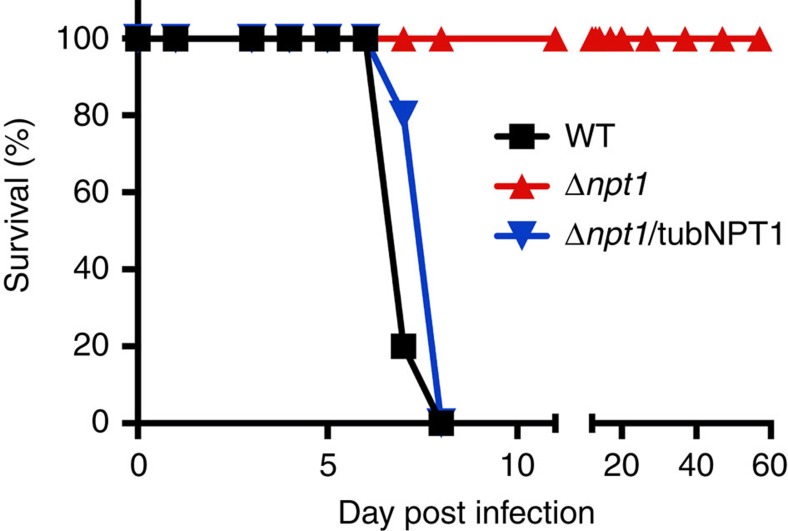
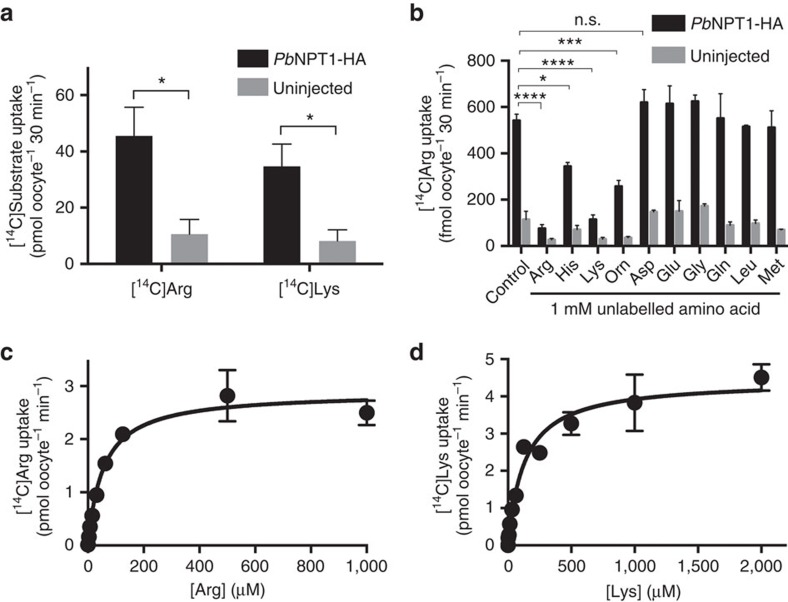
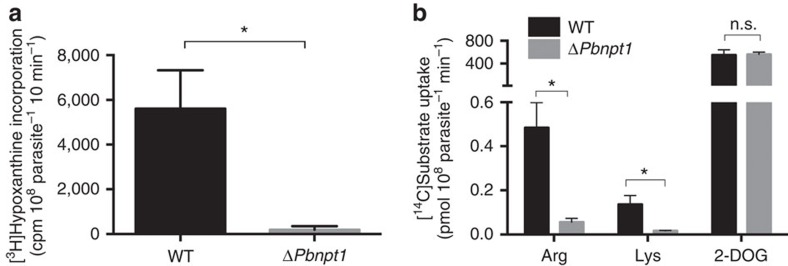
Apicomplexans are obligate intracellular parasites that scavenge essential nutrients from their hosts via transporter proteins on their plasma membrane. The identities of the transporters that mediate amino acid uptake into apicomplexans are unknown. Here we demonstrate that members of an apicomplexan-specific protein family-the Novel Putative Transporters (NPTs)-play key roles in the uptake of cationic amino acids. We show that an NPT from Toxoplasma gondii (TgNPT1) is a selective arginine transporter that is essential for parasite survival and virulence. We also demonstrate that a homologue of TgNPT1 from the malaria parasite Plasmodium berghei (PbNPT1), shown previously to be essential for the sexual gametocyte stage of the parasite, is a cationic amino acid transporter. This reveals a role for cationic amino acid scavenging in gametocyte biology. Our study demonstrates a critical role for amino acid transporters in the survival, virulence and life cycle progression of these parasites.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- van Dooren G. G. & Striepen B. The algal past and parasite present of the apicoplast. Annu. Rev. Microbiol. 67, 271–289 (2013). - PubMed
-
- Coppens I. Exploitation of auxotrophies and metabolic defects in Toxoplasma as therapeutic approaches. Int. J. Parasitol. 44, 109–120 (2014). - PubMed
-
- Carter N. S. et al.. Isolation and functional characterization of the PfNT1 nucleoside transporter gene from Plasmodium falciparum. J. Biol. Chem. 275, 10683–10691 (2000). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
