

Endoplasmic Reticulum (ER) Stress and Endocrine Disorders
- PMID: 28208663
- PMCID: PMC5343917
- DOI: 10.3390/ijms18020382
Endoplasmic Reticulum (ER) Stress and Endocrine Disorders
Abstract
The endoplasmic reticulum (ER) is the organelle where secretory and membrane proteins are synthesized and folded. Unfolded proteins that are retained within the ER can cause ER stress. Eukaryotic cells have a defense system called the "unfolded protein response" (UPR), which protects cells from ER stress. Cells undergo apoptosis when ER stress exceeds the capacity of the UPR, which has been revealed to cause human diseases. Although neurodegenerative diseases are well-known ER stress-related diseases, it has been discovered that endocrine diseases are also related to ER stress. In this review, we focus on ER stress-related human endocrine disorders. In addition to diabetes mellitus, which is well characterized, several relatively rare genetic disorders such as familial neurohypophyseal diabetes insipidus (FNDI), Wolfram syndrome, and isolated growth hormone deficiency type II (IGHD2) are discussed in this article.
Keywords: PKR-like endoplasmic reticulum kinase (PERK); activating transcription factor 6 (ATF6); chemical chaperone; endocrine disorder; endoplasmic reticulum stress; inositol requirement 1 (IRE1); old astrocyte specifically induced substance (OASIS) family.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
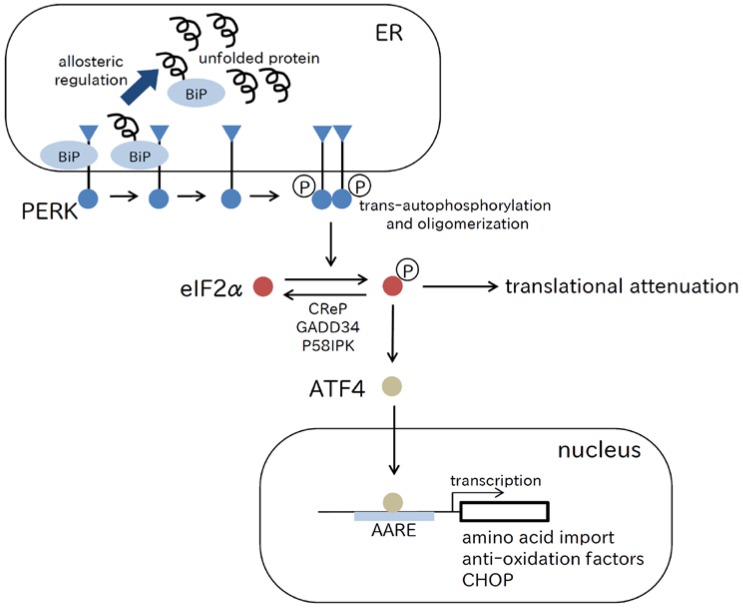
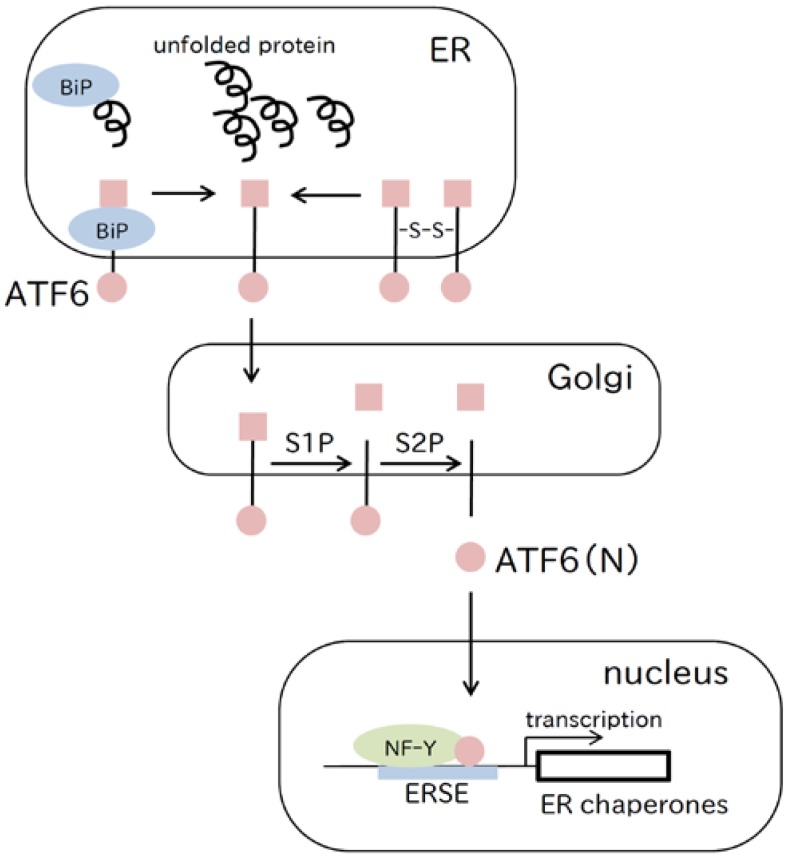
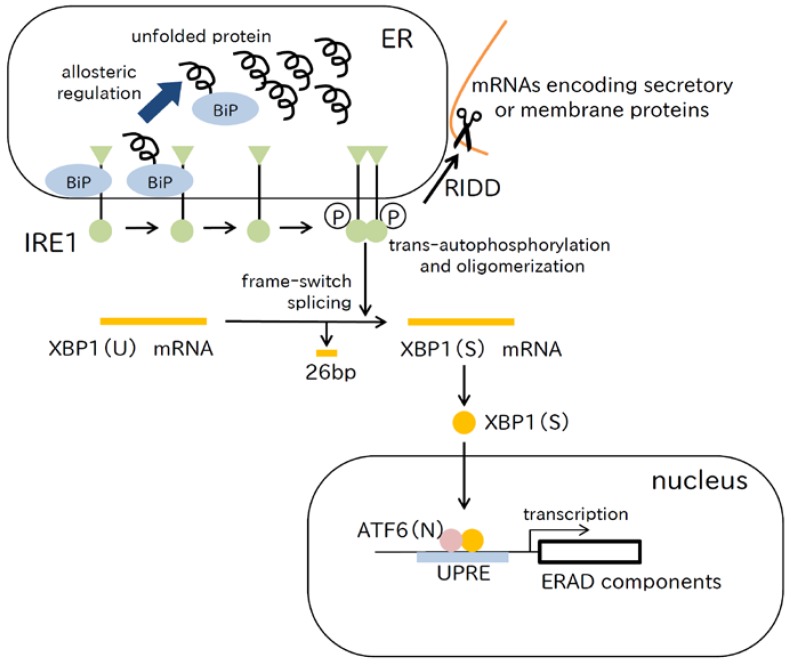
References
-
- Shimura H., Hattori N., Kubo S., Mizuno Y., Asakawa S., Minoshima S., Shimizu N., Iwai K., Chiba T., Tanaka K., et al. Familial Parkinson disease gene product, parkin, is a ubiquitin-protein ligase. Nat. Genet. 2000;25:302–305. - PubMed
-
- Schaffar G., Breuer P., Boteva R., Behrends C., Tzvetkov N., Strippel N., Sakahira H., Siegers K., Hayer-Hartl M., Hartl F.U. Cellular toxicity of polyglutamine expansion proteins: Mechanism of transcription factor deactivation. Mol. Cell. 2004;15:95–105. doi: 10.1016/j.molcel.2004.06.029. - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
