

Evaluation of the contribution of the transmembrane region to the ectodomain conformation of the human immunodeficiency virus (HIV-1) envelope glycoprotein
- PMID: 28209172
- PMCID: PMC5314615
- DOI: 10.1186/s12985-017-0704-x
Evaluation of the contribution of the transmembrane region to the ectodomain conformation of the human immunodeficiency virus (HIV-1) envelope glycoprotein
Abstract
Background: The human immunodeficiency virus (HIV-1) envelope glycoprotein (Env), a Type 1 transmembrane protein, assembles into a trimeric spike complex that mediates virus entry into host cells. The high potential energy of the metastable, unliganded Env trimer is maintained by multiple non-covalent contacts among the gp120 exterior and gp41 transmembrane Env subunits. Structural studies suggest that the gp41 transmembrane region forms a left-handed coiled coil that contributes to the Env trimer interprotomer contacts. Here we evaluate the contribution of the gp41 transmembrane region to the folding and stability of Env trimers.
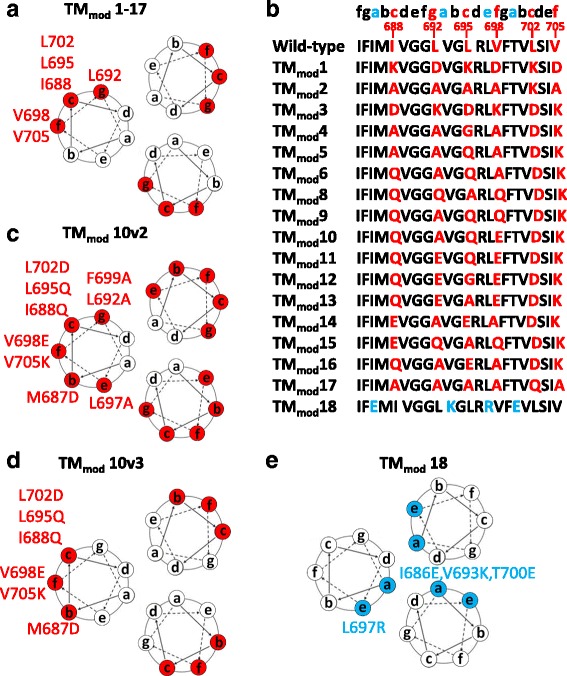
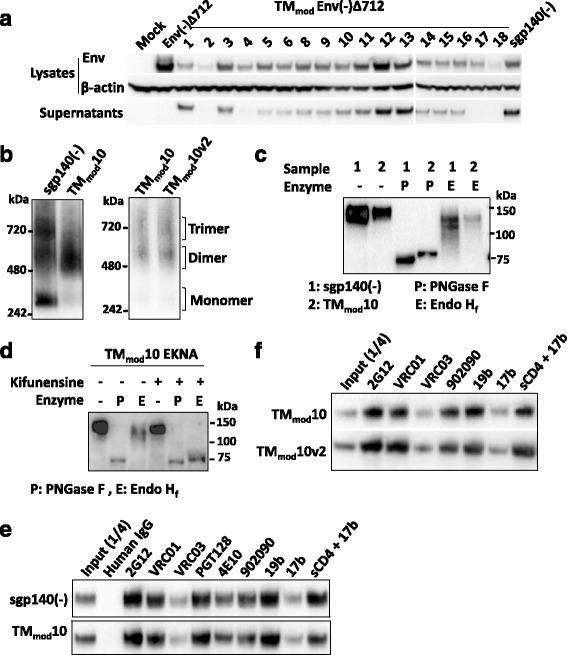
Methods: Multiple polar/charged amino acid residues, which hypothetically disrupt the stop-transfer signal, were introduced in the proposed lipid-interactive face of the transmembrane coiled coil, allowing release of soluble cleavage-negative Envs containing the modified transmembrane region (TMmod). We also examined effects of cleavage, the cytoplasmic tail and a C-terminal fibritin trimerization (FT) motif on oligomerization, antigenicity and functionality of soluble and membrane-bound Envs.
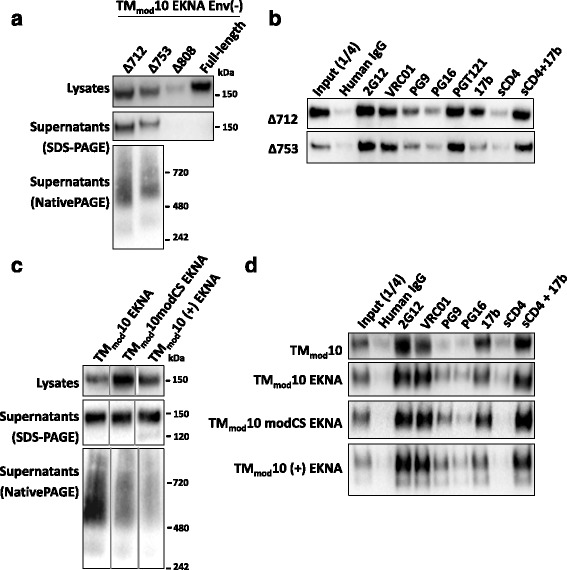
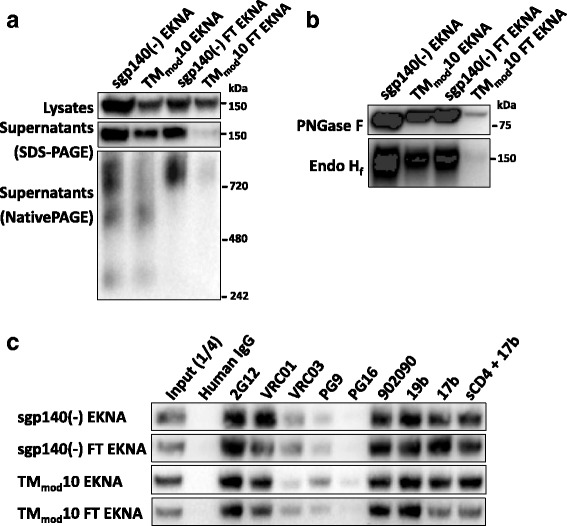
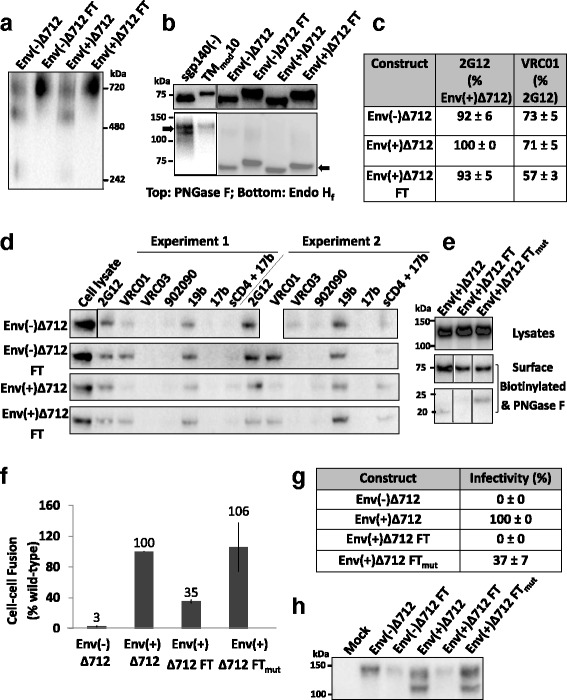
Results: The introduction of polar/charged amino acids into the transmembrane region resulted in the secretion of soluble Envs from the cell. However, these TMmod Envs primarily formed dimers. By contrast, control cleavage-negative sgp140 Envs lacking the transmembrane region formed soluble trimers, dimers and monomers. TMmod and sgp140 trimers were stabilized by the addition of a C-terminal FT sequence, but still exhibited carbohydrate and antigenic signatures of a flexible ectodomain structure. On the other hand, detergent-solubilized cleaved and uncleaved Envs isolated from the membranes of expressing cells exhibited "tighter" ectodomain structures, based on carbohydrate modifications. These trimers were found to be unstable in detergent solutions, but could be stabilized by the addition of a C-terminal FT moiety. The C-terminal FT domain decreased Env cleavage and syncytium-forming ability by approximately three-fold; alteration of the FT trimerization interface restored Env cleavage and syncytium formation to near-wild-type levels.
Conclusion: The modified transmembrane region was not conducive to trimerization of soluble Envs. However, for HIV-1 Env ectodomains that are minimally modified, membrane-anchored Envs exhibit the most native structures and can be stabilized by appropriately positioned FT domains.
Keywords: Ectodomain; Fibritin –Trimer; HIV-1 Env; Stabilize; Transmembrane region; gp41.
Figures
Similar articles
-
Conformational Differences between Functional Human Immunodeficiency Virus Envelope Glycoprotein Trimers and Stabilized Soluble Trimers.J Virol. 2019 Jan 17;93(3):e01709-18. doi: 10.1128/JVI.01709-18. Print 2019 Feb 1. J Virol. 2019. PMID: 30429345 Free PMC article.
-
Effects of the I559P gp41 change on the conformation and function of the human immunodeficiency virus (HIV-1) membrane envelope glycoprotein trimer.PLoS One. 2015 Apr 7;10(4):e0122111. doi: 10.1371/journal.pone.0122111. eCollection 2015. PLoS One. 2015. PMID: 25849367 Free PMC article.
-
Comparison of Uncleaved and Mature Human Immunodeficiency Virus Membrane Envelope Glycoprotein Trimers.J Virol. 2018 May 29;92(12):e00277-18. doi: 10.1128/JVI.00277-18. Print 2018 Jun 15. J Virol. 2018. PMID: 29618643 Free PMC article.
-
HIV-1 envelope glycoprotein structure.Curr Opin Struct Biol. 2013 Apr;23(2):268-76. doi: 10.1016/j.sbi.2013.03.007. Epub 2013 Apr 18. Curr Opin Struct Biol. 2013. PMID: 23602427 Free PMC article. Review.
-
The HIV Env-mediated fusion reaction.Biochim Biophys Acta. 2003 Jul 11;1614(1):36-50. doi: 10.1016/s0005-2736(03)00161-5. Biochim Biophys Acta. 2003. PMID: 12873764 Review.
Cited by
-
Characterization of HIV-1 Nucleoside-Modified mRNA Vaccines in Rabbits and Rhesus Macaques.Mol Ther Nucleic Acids. 2019 Apr 15;15:36-47. doi: 10.1016/j.omtn.2019.03.003. Epub 2019 Mar 21. Mol Ther Nucleic Acids. 2019. PMID: 30974332 Free PMC article.
-
HIV-1-based Virus-like Particles that Morphologically Resemble Mature, Infectious HIV-1 Virions.Viruses. 2019 Jun 2;11(6):507. doi: 10.3390/v11060507. Viruses. 2019. PMID: 31159488 Free PMC article.
-
Strain-Dependent Activation and Inhibition of Human Immunodeficiency Virus Entry by a Specific PF-68742 Stereoisomer.J Virol. 2019 Oct 15;93(21):e01197-19. doi: 10.1128/JVI.01197-19. Print 2019 Nov 1. J Virol. 2019. PMID: 31391272 Free PMC article.
-
Conformational Differences between Functional Human Immunodeficiency Virus Envelope Glycoprotein Trimers and Stabilized Soluble Trimers.J Virol. 2019 Jan 17;93(3):e01709-18. doi: 10.1128/JVI.01709-18. Print 2019 Feb 1. J Virol. 2019. PMID: 30429345 Free PMC article.
-
Effects of the SOS (A501C/T605C) and DS (I201C/A433C) Disulfide Bonds on HIV-1 Membrane Envelope Glycoprotein Conformation and Function.J Virol. 2019 May 29;93(12):e00304-19. doi: 10.1128/JVI.00304-19. Print 2019 Jun 15. J Virol. 2019. PMID: 30944182 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
