

Small Molecules That Sabotage Bacterial Virulence
- PMID: 28209403
- PMCID: PMC5679398
- DOI: 10.1016/j.tips.2017.01.004
Small Molecules That Sabotage Bacterial Virulence
Abstract
The continued rise of antibiotic-resistant bacterial infections has motivated alternative strategies for target discovery and treatment of infections. Antivirulence therapies function through inhibition of in vivo required virulence factors to disarm the pathogen instead of directly targeting viability or growth. This approach to treating bacteria-mediated diseases may have advantages over traditional antibiotics because it targets factors specific for pathogenesis, potentially reducing selection for resistance and limiting collateral damage to the resident microbiota. This review examines vulnerable molecular mechanisms used by bacteria to cause disease and the antivirulence compounds that sabotage these virulence pathways. By expanding the study of antimicrobial targets beyond those that are essential for growth, antivirulence strategies offer new and innovative opportunities to combat infectious diseases.
Keywords: antivirulence therapies; bacterial pathogenesis.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Conflict of interest statement
The authors have no competing interests in the presented work.
Figures
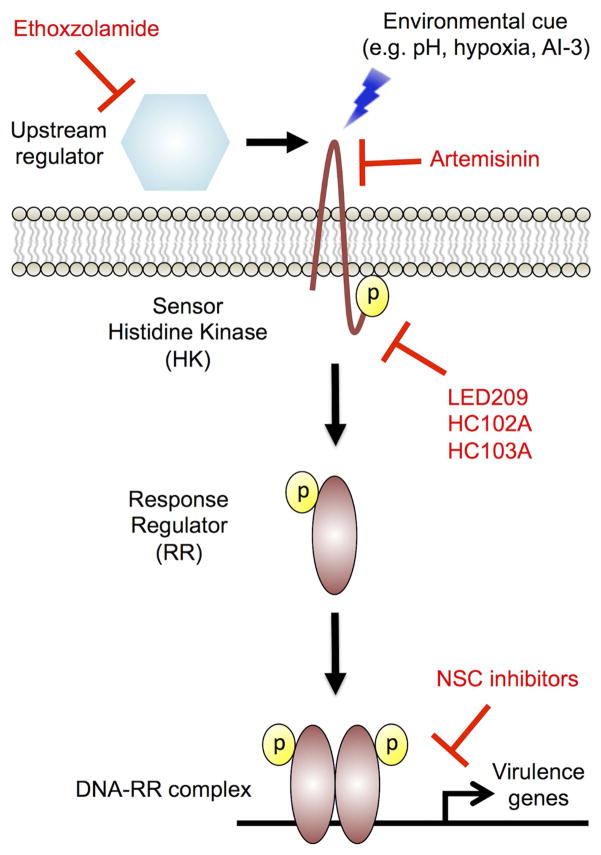
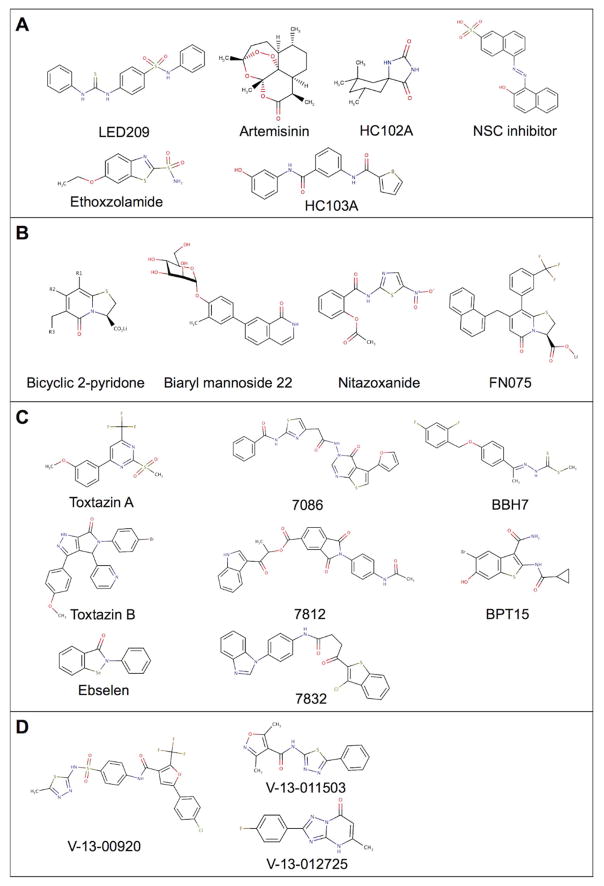
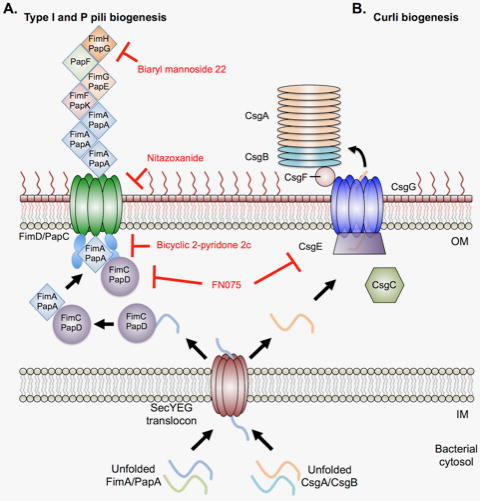
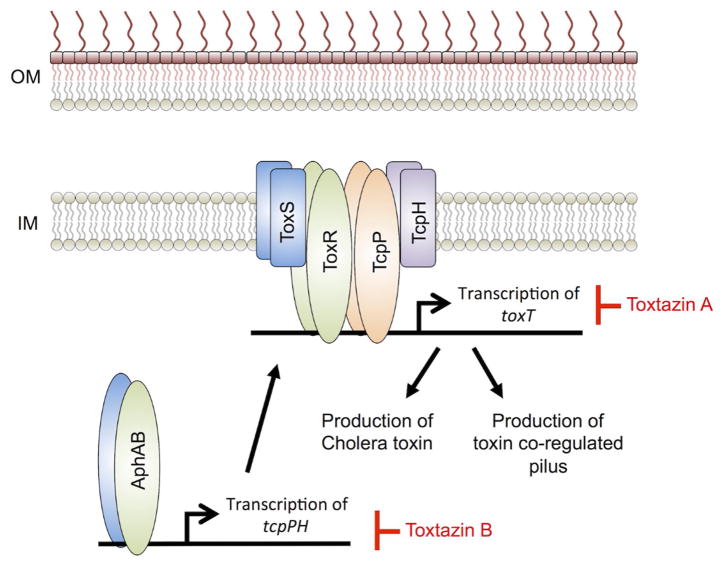
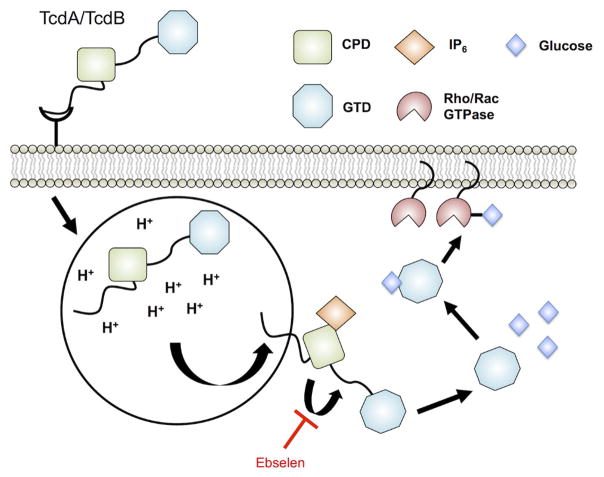
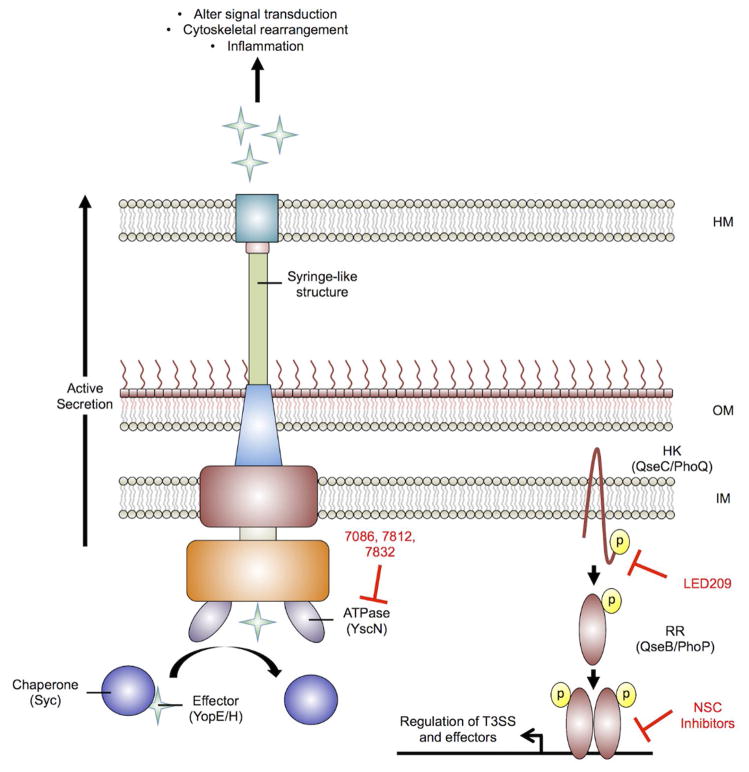
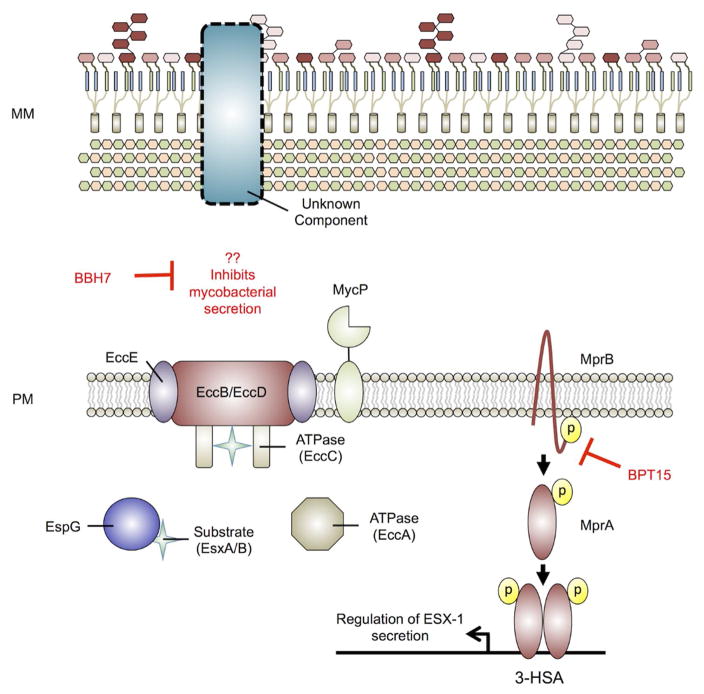
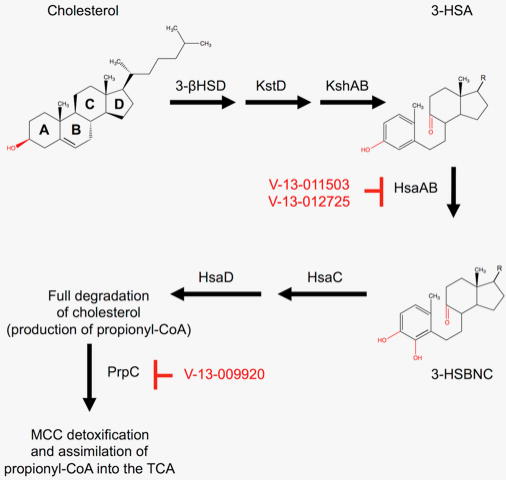
References
-
- D’Costa VM, et al. Sampling the antibiotic resistome. Science. 2006;311:374–377. - PubMed
-
- CDC. Antibiotic Resistance Threats in the United States, 2013. Centers for Disease Control and Prevention; Atlanta, GA: 2013.
-
- CDC. Antimicrobial Resistance Posing Growing Health Threat. Center for Disease Control and Prevention; Atlanta, GA: 2011.
-
- Payne DJ, et al. Drugs for bad bugs: confronting the challenges of antibacterial discovery. Nature reviews Drug discovery. 2007;6:29–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
