

Molecular Interplay between the Dimer Interface and the Substrate-Binding Site of Human Peptidylarginine Deiminase 4
- PMID: 28209966
- PMCID: PMC5314407
- DOI: 10.1038/srep42662
Molecular Interplay between the Dimer Interface and the Substrate-Binding Site of Human Peptidylarginine Deiminase 4
Abstract
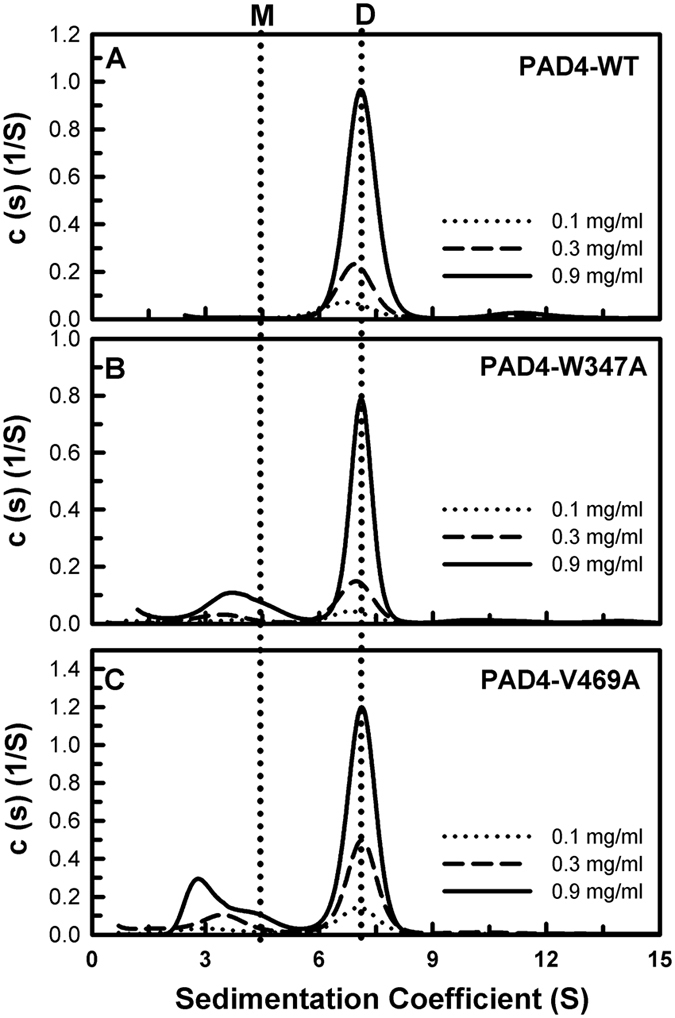
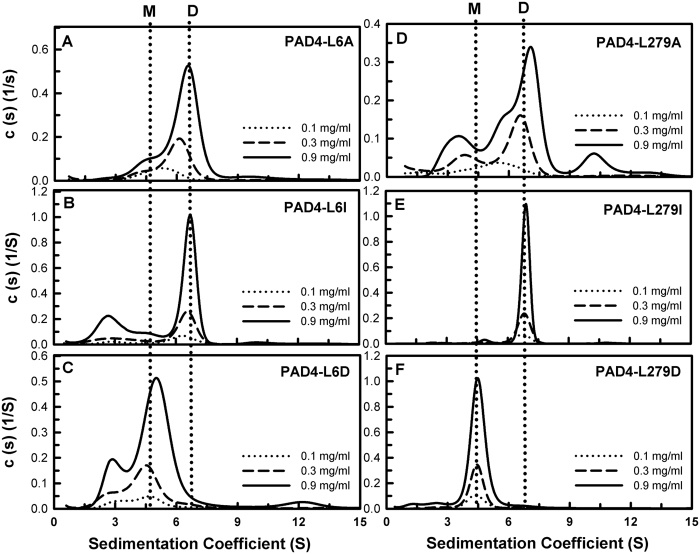
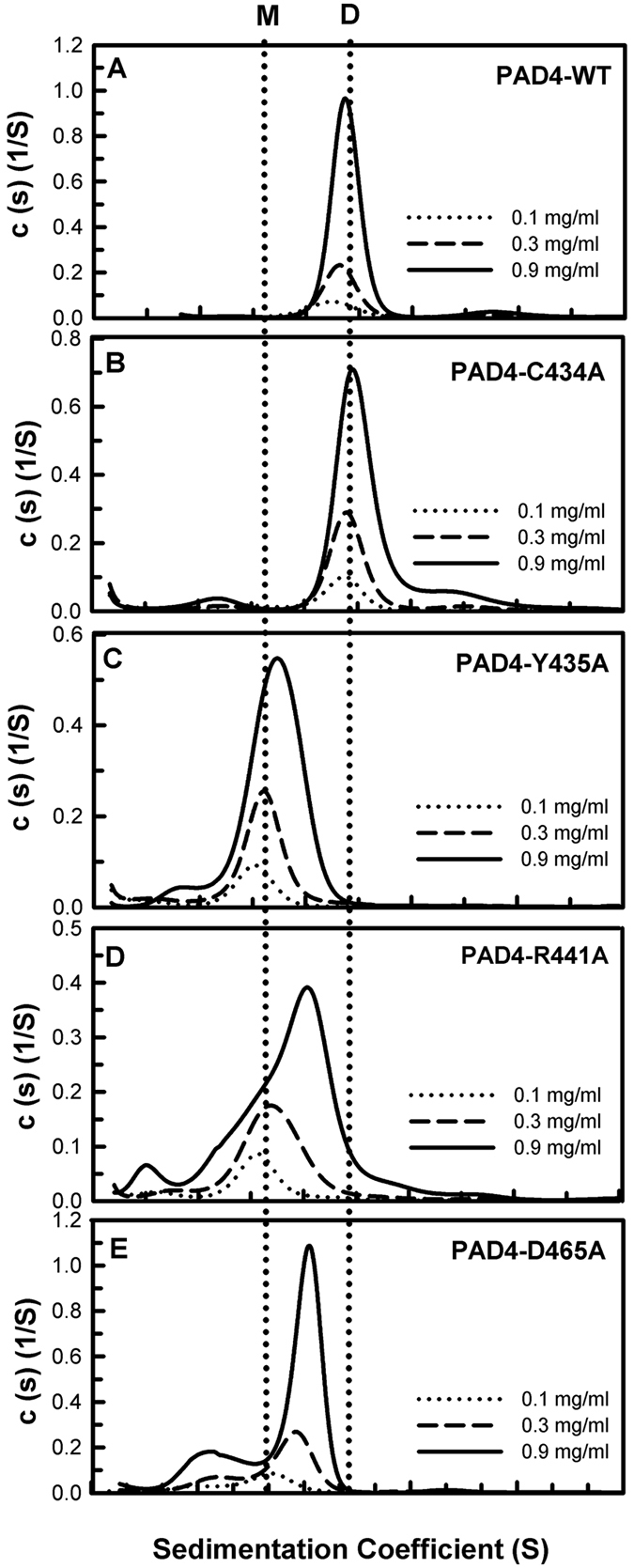
Our previous studies suggest that the fully active form of Peptidylarginine deiminase 4 (PAD4) should be a dimer and not a monomer. This paper provides a plausible mechanism for the control of PAD4 catalysis by molecular interplay between its dimer-interface loop (I-loop) and its substrate-binding loop (S-loop). Mutagenesis studies revealed that two hydrophobic residues, W347 and V469, are critical for substrate binding at the active site; mutating these two residues led to a severe reduction in the catalytic activity. We also identified several hydrophobic amino acid residues (L6, L279 and V283) at the dimer interface. Ultracentrifugation analysis revealed that interruption of the hydrophobicity of this region decreases dimer formation and, consequently, enzyme activity. Molecular dynamic simulations and mutagenesis studies suggested that the dimer interface and the substrate-binding site of PAD4, which consist of the I-loop and the S-loop, respectively, are responsible for substrate binding and dimer stabilization. We identified five residues with crucial roles in PAD4 catalysis and dimerization: Y435 and R441 in the I-loop, D465 and V469 in the S-loop, and W548, which stabilizes the I-loop via van der Waals interactions with C434 and Y435. The molecular interplay between the S-loop and the I-loop is crucial for PAD4 catalysis.
Conflict of interest statement
The authors declare no competing financial interests.
Figures




Similar articles
-
Catalytic activity of human guanylate-binding protein 1 coupled to the release of structural restraints imposed by the C-terminal domain.FEBS J. 2021 Jan;288(2):582-599. doi: 10.1111/febs.15348. Epub 2020 May 19. FEBS J. 2021. PMID: 32352209
-
Selectively Modulating Conformational States of USP7 Catalytic Domain for Activation.Structure. 2018 Jan 2;26(1):72-84.e7. doi: 10.1016/j.str.2017.11.010. Epub 2017 Dec 14. Structure. 2018. PMID: 29249604
-
Differential lipid binding of vinculin isoforms promotes quasi-equivalent dimerization.Proc Natl Acad Sci U S A. 2016 Aug 23;113(34):9539-44. doi: 10.1073/pnas.1600702113. Epub 2016 Aug 8. Proc Natl Acad Sci U S A. 2016. PMID: 27503891 Free PMC article.
-
Structural and molecular dynamics studies of a C1-oxidizing lytic polysaccharide monooxygenase from Heterobasidion irregulare reveal amino acids important for substrate recognition.FEBS J. 2018 Jun;285(12):2225-2242. doi: 10.1111/febs.14472. Epub 2018 Apr 24. FEBS J. 2018. PMID: 29660793
-
Structural basis for catalysis at the membrane-water interface.Biochim Biophys Acta Mol Cell Biol Lipids. 2017 Nov;1862(11):1368-1385. doi: 10.1016/j.bbalip.2016.11.011. Epub 2016 Nov 30. Biochim Biophys Acta Mol Cell Biol Lipids. 2017. PMID: 27913292 Free PMC article. Review.
Cited by
-
Peptidylarginine deiminase enzymes and citrullinated proteins in female reproductive physiology and associated diseases†.Biol Reprod. 2022 Dec 10;107(6):1395-1410. doi: 10.1093/biolre/ioac173. Biol Reprod. 2022. PMID: 36087287 Free PMC article. Review.
-
Antibody discovery identifies regulatory mechanisms of protein arginine deiminase 4.Nat Chem Biol. 2024 Jun;20(6):742-750. doi: 10.1038/s41589-023-01535-8. Epub 2024 Feb 2. Nat Chem Biol. 2024. PMID: 38308046 Free PMC article.
-
Probing the Roles of Calcium-Binding Sites during the Folding of Human Peptidylarginine Deiminase 4.Sci Rep. 2017 May 25;7(1):2429. doi: 10.1038/s41598-017-02677-1. Sci Rep. 2017. PMID: 28546558 Free PMC article.
-
Crystal structure of human peptidylarginine deiminase type VI (PAD6) provides insights into its inactivity.IUCrJ. 2024 May 1;11(Pt 3):395-404. doi: 10.1107/S2052252524002549. IUCrJ. 2024. PMID: 38656308 Free PMC article.
-
The virtues and vices of protein citrullination.R Soc Open Sci. 2022 Jun 8;9(6):220125. doi: 10.1098/rsos.220125. eCollection 2022 Jun. R Soc Open Sci. 2022. PMID: 35706669 Free PMC article. Review.
References
-
- Gyorgy B., Toth E., Tarcsa E., Falus A. & Buzas E. I. Citrullination: a posttranslational modification in health and disease. Int. J. Biochem. Cell Biol. 38, 1662–1677 (2006). - PubMed
-
- Rogers G., Winter B., McLaughlan C., Powell B. & Nesci T. Peptidylarginine deiminase of the hair follicle: characterization, localization, and function in keratinizing tissues. J. Invest. Dermatol. 108, 700–707 (1997). - PubMed
-
- Boggs J. M. et al.. Highly deiminated isoform of myelin basic protein from multiple sclerosis brain causes fragmentation of lipid vesicles. J. Neurosci. Res. 57, 529–535 (1999). - PubMed
-
- Vossenaar E. R., Zendman A. J., van Venrooij W. J. & Pruijn G. J. PAD, a growing family of citrullinating enzymes: genes, features and involvement in disease. BioEssays 25, 1106–1118 (2003). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
