

OTX2 Activity at Distal Regulatory Elements Shapes the Chromatin Landscape of Group 3 Medulloblastoma
- PMID: 28213356
- PMCID: PMC5340587
- DOI: 10.1158/2159-8290.CD-16-0844
OTX2 Activity at Distal Regulatory Elements Shapes the Chromatin Landscape of Group 3 Medulloblastoma
Abstract
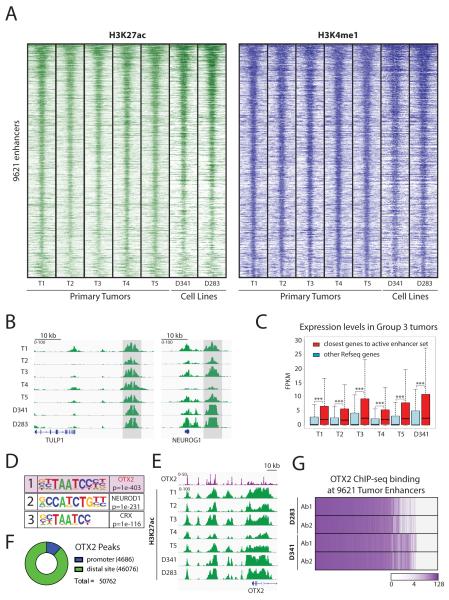
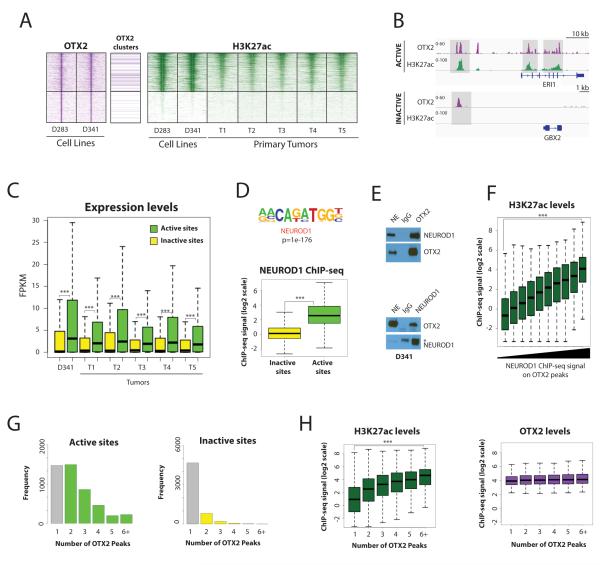
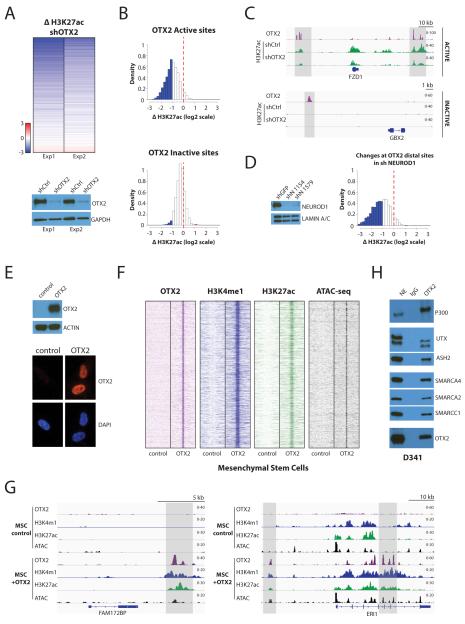
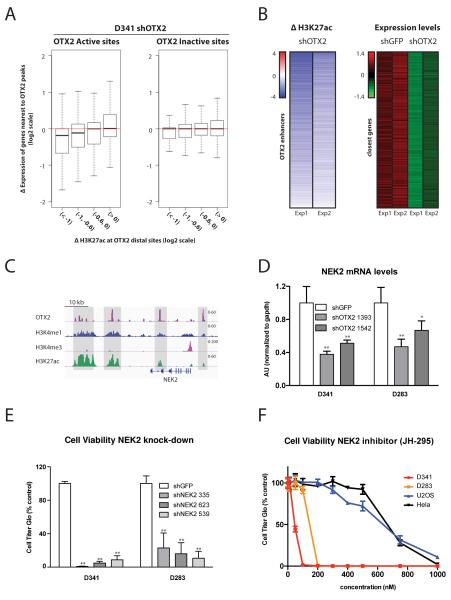
Medulloblastoma is the most frequent malignant pediatric brain tumor and is divided into at least four subgroups known as WNT, SHH, Group 3, and Group 4. Here, we characterized gene regulation mechanisms in the most aggressive subtype, Group 3 tumors, through genome-wide chromatin and expression profiling. Our results show that most active distal sites in these tumors are occupied by the transcription factor OTX2. Highly active OTX2-bound enhancers are often arranged as clusters of adjacent peaks and are also bound by the transcription factor NEUROD1. These sites are responsive to OTX2 and NEUROD1 knockdown and could also be generated de novo upon ectopic OTX2 expression in primary cells, showing that OTX2 cooperates with NEUROD1 and plays a major role in maintaining and possibly establishing regulatory elements as a pioneer factor. Among OTX2 target genes, we identified the kinase NEK2, whose knockdown and pharmacologic inhibition decreased cell viability. Our studies thus show that OTX2 controls the regulatory landscape of Group 3 medulloblastoma through cooperative activity at enhancer elements and contributes to the expression of critical target genes.Significance: The gene regulation mechanisms that drive medulloblastoma are not well understood. Using chromatin profiling, we find that the transcription factor OTX2 acts as a pioneer factor and, in cooperation with NEUROD1, controls the Group 3 medulloblastoma active enhancer landscape. OTX2 itself or its target genes, including the mitotic kinase NEK2, represent attractive targets for future therapies. Cancer Discov; 7(3); 288-301. ©2017 AACR.This article is highlighted in the In This Issue feature, p. 235.
©2017 American Association for Cancer Research.
Figures
References
-
- Thompson MC, Fuller C, Hogg TL, Dalton J, Finkelstein D, Lau CC, et al. Genomics identifies medulloblastoma subgroups that are enriched for specific genetic alterations. J Clin Oncol. 2006;24:1924–31. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
