

HSP90 Shapes the Consequences of Human Genetic Variation
- PMID: 28215707
- PMCID: PMC5325763
- DOI: 10.1016/j.cell.2017.01.023
HSP90 Shapes the Consequences of Human Genetic Variation
Abstract
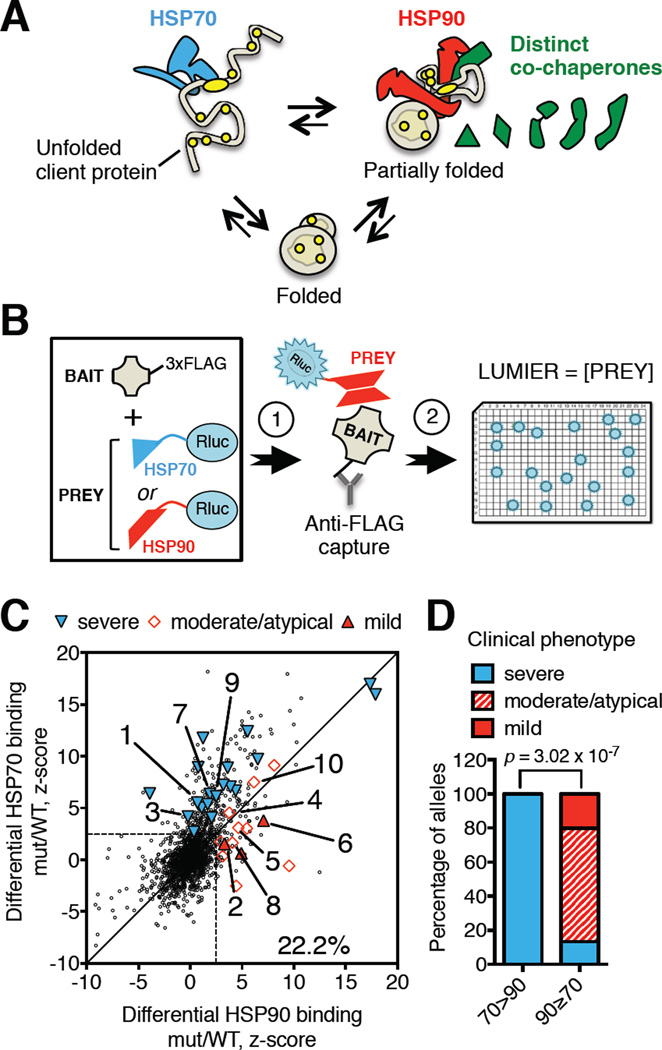
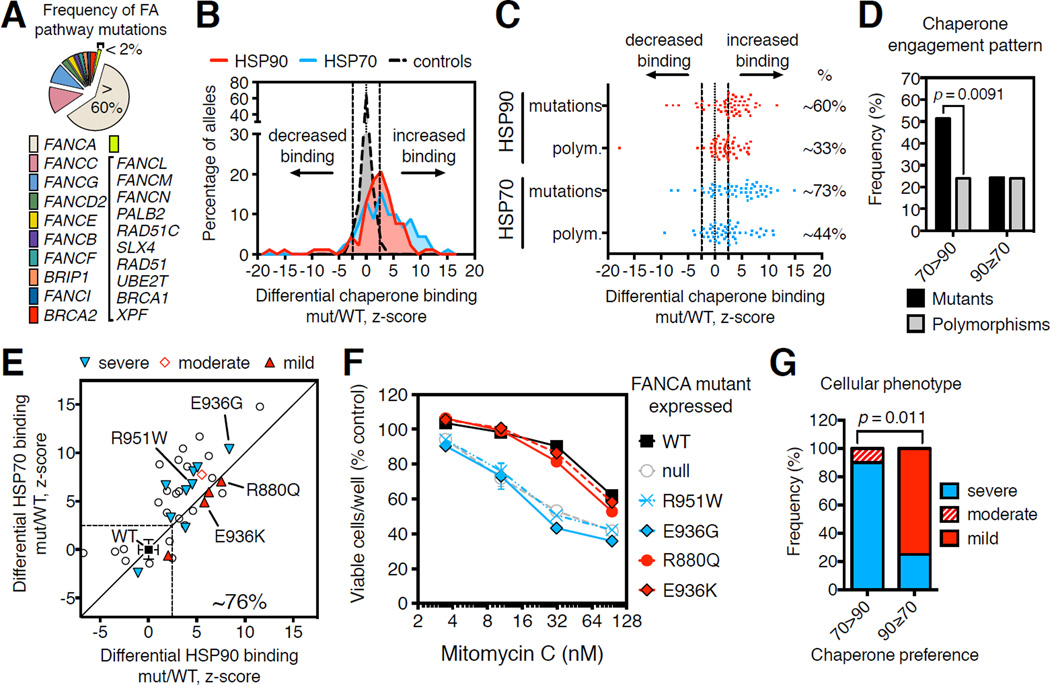
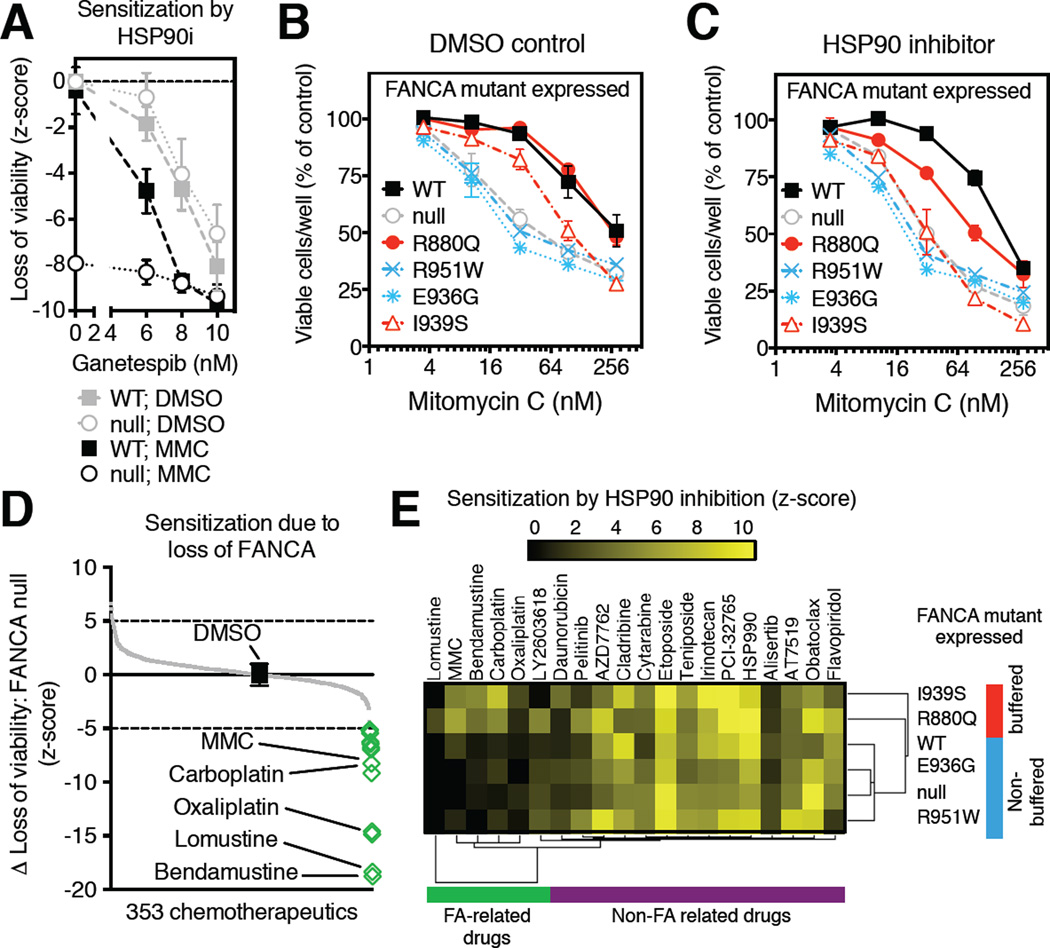
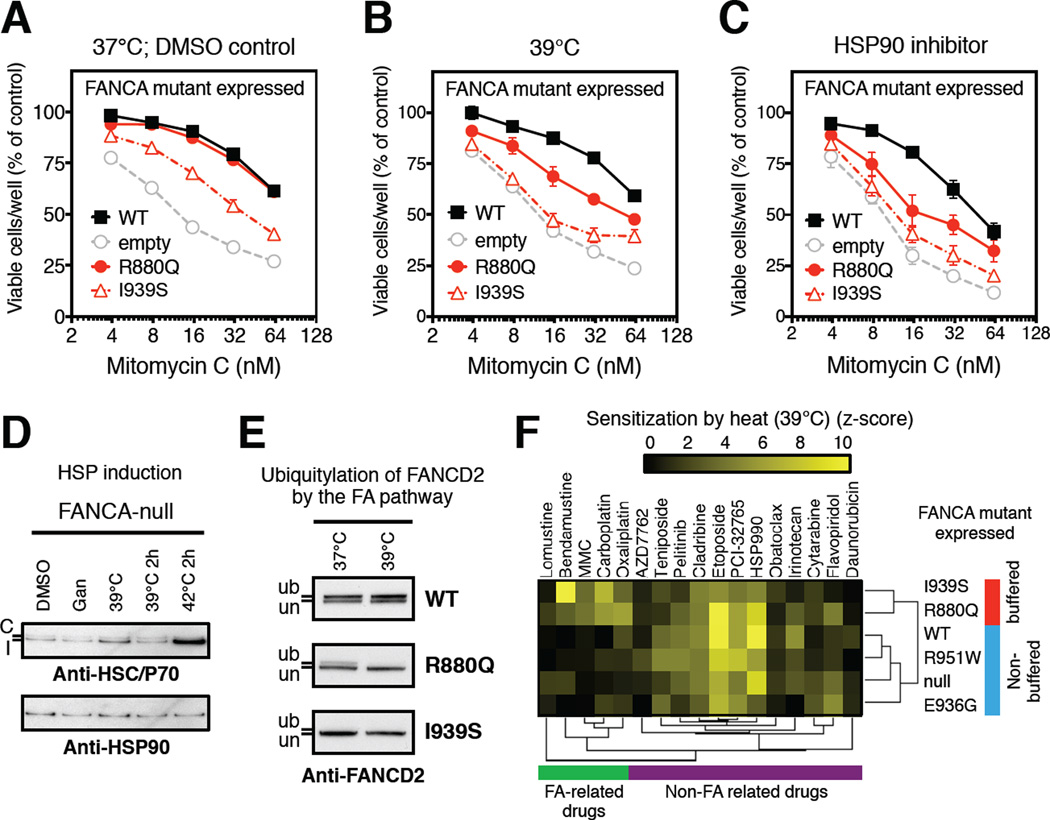
HSP90 acts as a protein-folding buffer that shapes the manifestations of genetic variation in model organisms. Whether HSP90 influences the consequences of mutations in humans, potentially modifying the clinical course of genetic diseases, remains unknown. By mining data for >1,500 disease-causing mutants, we found a strong correlation between reduced phenotypic severity and a dominant (HSP90 ≥ HSP70) increase in mutant engagement by HSP90. Examining the cancer predisposition syndrome Fanconi anemia in depth revealed that mutant FANCA proteins engaged predominantly by HSP70 had severely compromised function. In contrast, the function of less severe mutants was preserved by a dominant increase in HSP90 binding. Reducing HSP90's buffering capacity with inhibitors or febrile temperatures destabilized HSP90-buffered mutants, exacerbating FA-related chemosensitivities. Strikingly, a compensatory FANCA somatic mutation from an "experiment of nature" in monozygotic twins both prevented anemia and reduced HSP90 binding. These findings provide one plausible mechanism for the variable expressivity and environmental sensitivity of genetic diseases.
Keywords: Fanconi anemia; HSP70; HSP90 buffering; cancer; gene-environment interaction.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Genetic variation: HSP90 shapes human genetic variation.Nat Rev Genet. 2017 Mar 15;18(4):210-211. doi: 10.1038/nrg.2017.16. Nat Rev Genet. 2017. PMID: 28293031 No abstract available.
-
Molecular genetics: Chaperone protein gets personal.Nature. 2017 May 4;545(7652):36-37. doi: 10.1038/nature22487. Epub 2017 Apr 19. Nature. 2017. PMID: 28424517 No abstract available.
References
-
- Adachi D, Oda T, Yagasaki H, Nakasato K, Taniguchi T, D’Andrea AD, Asano S, Yamashita T. Heterogeneous activation of the Fanconi anemia pathway by patient-derived FANCA mutants. Human molecular genetics. 2002;11:3125–3134. - PubMed
-
- Barrios-Rodiles M, Brown KR, Ozdamar B, Bose R, Liu Z, Donovan RS, Shinjo F, Liu Y, Dembowy J, Taylor IW, et al. High-throughput mapping of a dynamic signaling network in mammalian cells. Science. 2005;307:1621–1625. - PubMed
-
- Biesecker LG, Spinner NB. A genomic view of mosaicism and human disease. Nature reviews Genetics. 2013;14:307–320. - PubMed
-
- Burga A, Casanueva MO, Lehner B. Predicting mutation outcome from early stochastic variation in genetic interaction partners. Nature. 2011;480:250–253. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
