

Converting Adult Pancreatic Islet α Cells into β Cells by Targeting Both Dnmt1 and Arx
- PMID: 28215845
- PMCID: PMC5358097
- DOI: 10.1016/j.cmet.2017.01.009
Converting Adult Pancreatic Islet α Cells into β Cells by Targeting Both Dnmt1 and Arx
Abstract
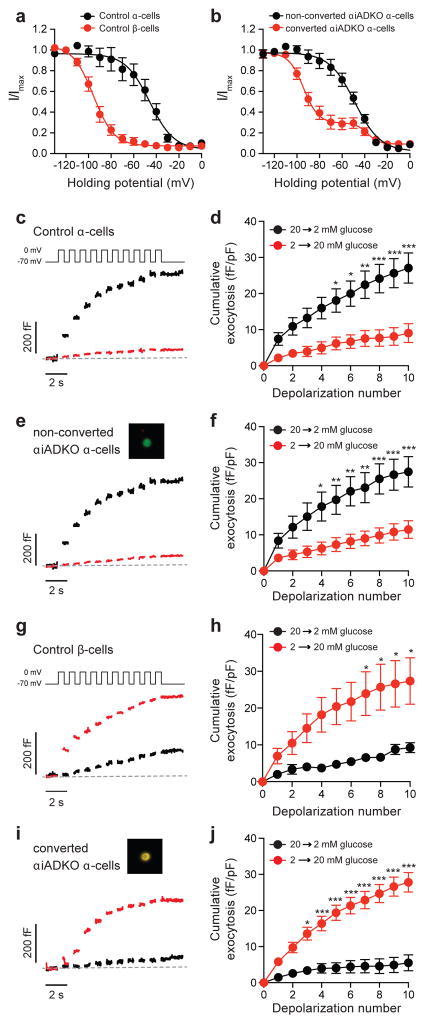
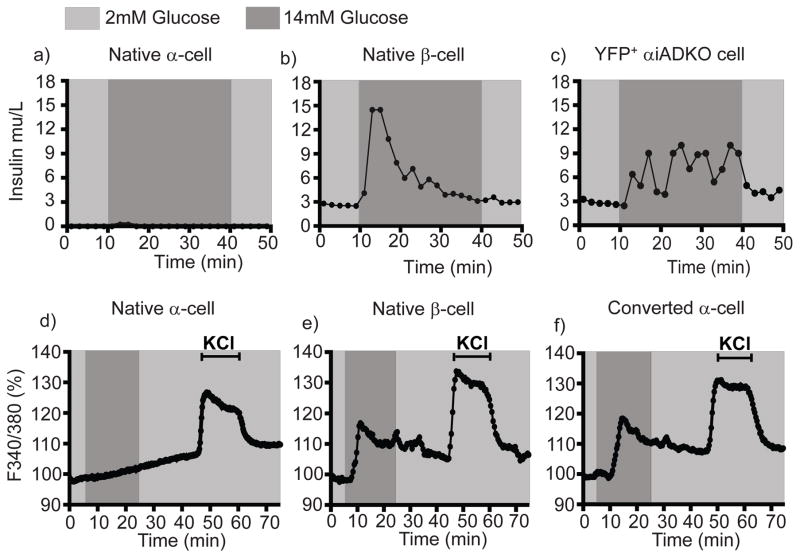
Insulin-producing pancreatic β cells in mice can slowly regenerate from glucagon-producing α cells in settings like β cell loss, but the basis of this conversion is unknown. Moreover, it remains unclear if this intra-islet cell conversion is relevant to diseases like type 1 diabetes (T1D). We show that the α cell regulators Aristaless-related homeobox (Arx) and DNA methyltransferase 1 (Dnmt1) maintain α cell identity in mice. Within 3 months of Dnmt1 and Arx loss, lineage tracing and single-cell RNA sequencing revealed extensive α cell conversion into progeny resembling native β cells. Physiological studies demonstrated that converted α cells acquire hallmark β cell electrophysiology and show glucose-stimulated insulin secretion. In T1D patients, subsets of glucagon-expressing cells show loss of DNMT1 and ARX and produce insulin and other β cell factors, suggesting that DNMT1 and ARX maintain α cell identity in humans. Our work reveals pathways regulated by Arx and Dnmt1 that are sufficient for achieving targeted generation of β cells from adult pancreatic α cells.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
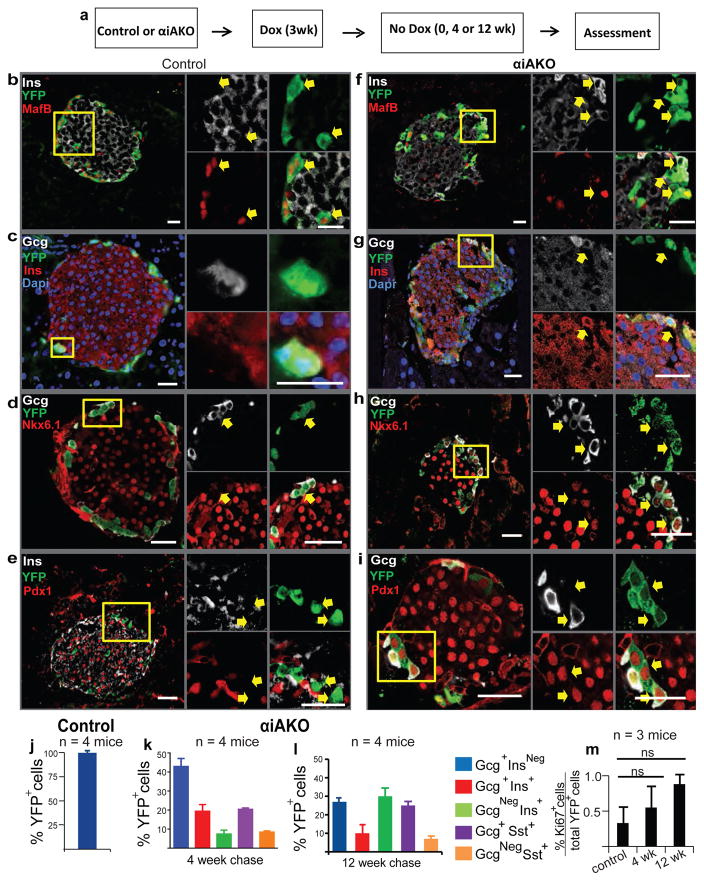
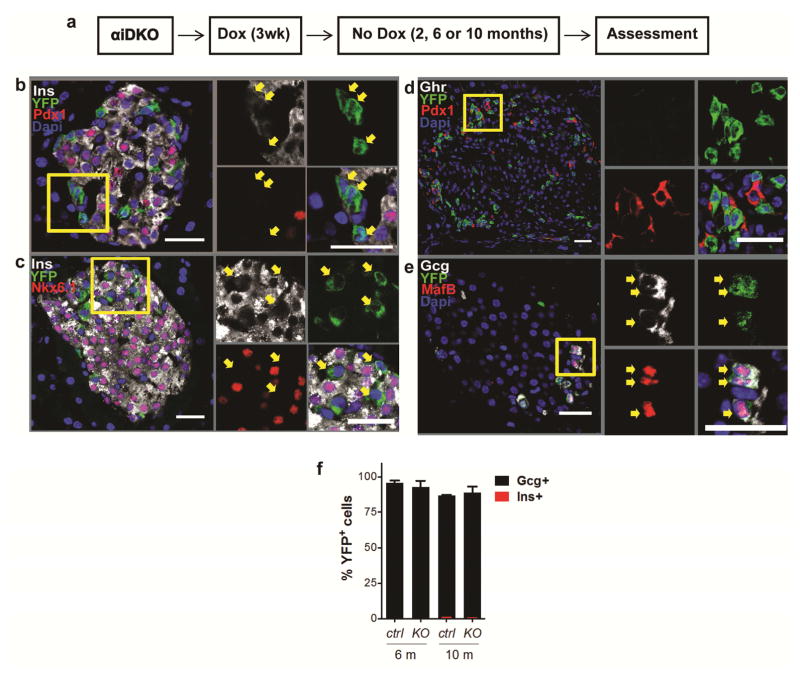
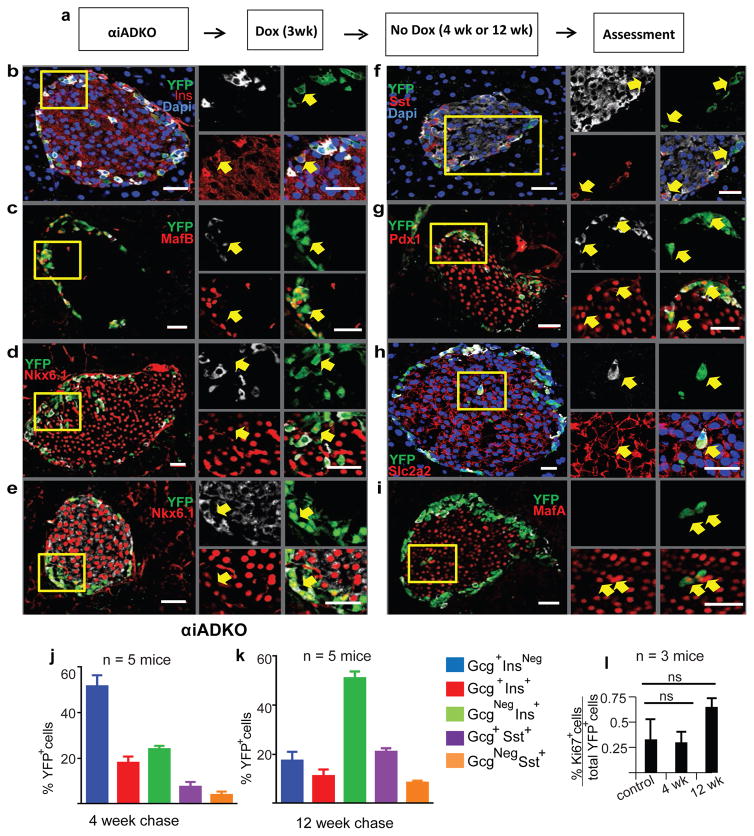
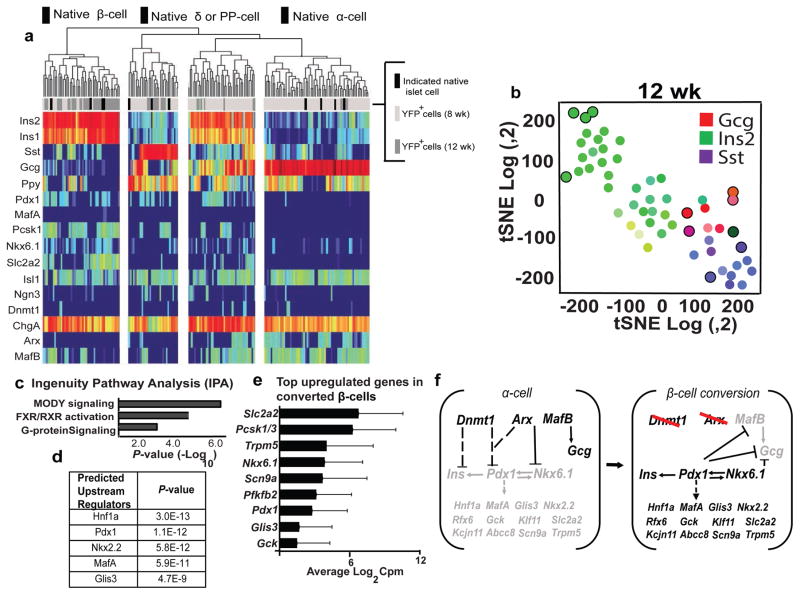
Comment in
-
Should I Stay or Should I Go: A Clash of α-Cell Identity.Cell Metab. 2017 Mar 7;25(3):488-490. doi: 10.1016/j.cmet.2017.02.014. Cell Metab. 2017. PMID: 28273469
References
-
- Adewola AF, Lee D, Harvat T, Mohammed J, Eddington DT, Oberholzer J, Wang Y. Microfluidic perifusion and imaging device for multi-parametric islet function assessment. Biomed Microdevices. 2010;12(3):409–17. - PubMed
-
- Arda HE, Li L, Tsai J, Torre EA, Rosli Y, Peiris H, Spitale RC, Dai C, Gu X, Qu K, Wang P, Wang J, Grompe M, Scharfmann R, Snyder MS, Bottino R, Powers AC, Chang HY, Kim SK. Age-dependent pancreatic gene regulation reveals mechanisms governing human β-cell function. Cell Metab. 2016;23(5):909–20. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
