

The β-barrel assembly machinery in motion
- PMID: 28216659
- PMCID: PMC5455337
- DOI: 10.1038/nrmicro.2016.191
The β-barrel assembly machinery in motion
Abstract
In Gram-negative bacteria, the biogenesis of β-barrel outer membrane proteins (OMPs) is mediated by the β-barrel assembly machinery (BAM) complex. During the past decade, structural and functional studies have collectively contributed to advancing our understanding of the structure and function of the BAM complex; however, the exact mechanism that is involved remains elusive. In this Progress article, we discuss recent structural studies that have revealed that the accessory proteins may regulate essential unprecedented conformational changes in the core component BamA during function. We also detail the mechanistic insights that have been gained from structural data, mutagenesis studies and molecular dynamics simulations, and explore two emerging models for the BAM-mediated biogenesis of OMPs in bacteria.
Conflict of interest statement
The authors declare no competing interests.
Figures
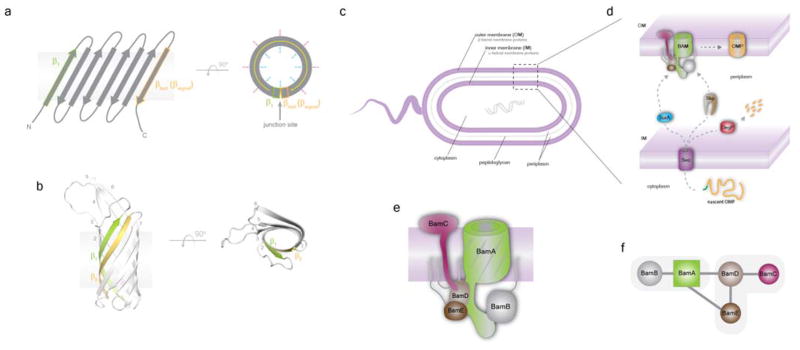
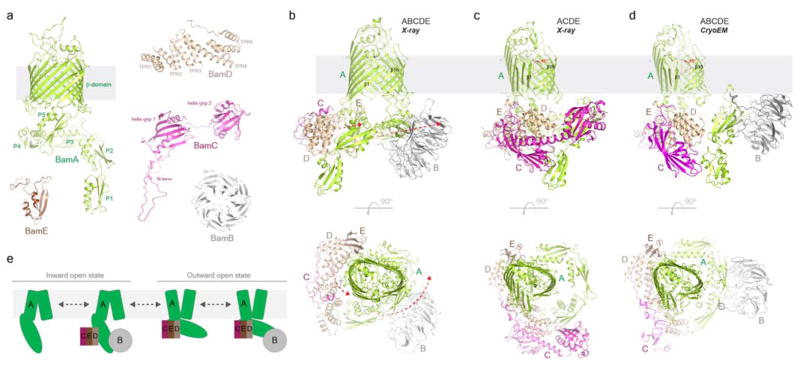

References
-
- Schulz GE. beta-Barrel membrane proteins. Curr Opin Struct Biol. 2000;10:443–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
