

Anaphase A: Disassembling Microtubules Move Chromosomes toward Spindle Poles
- PMID: 28218660
- PMCID: PMC5372008
- DOI: 10.3390/biology6010015
Anaphase A: Disassembling Microtubules Move Chromosomes toward Spindle Poles
Abstract
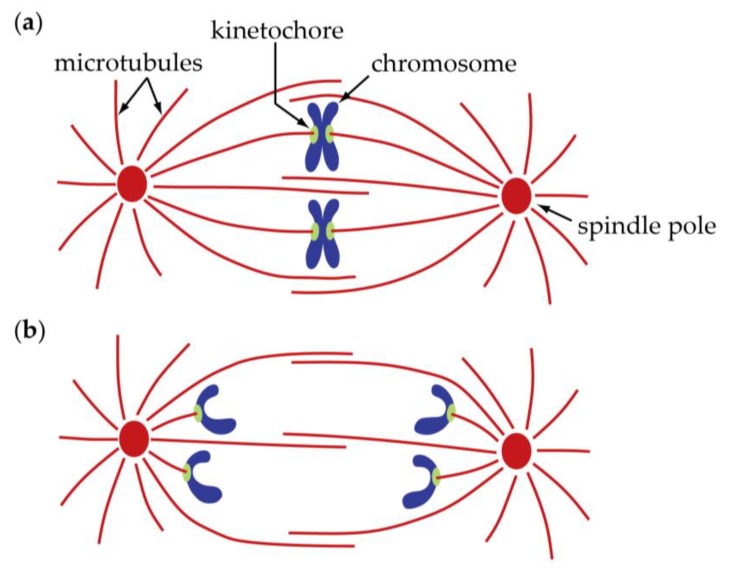
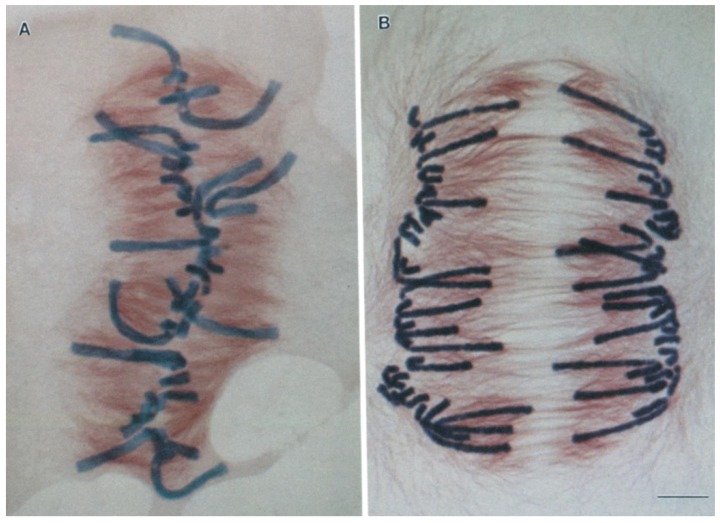
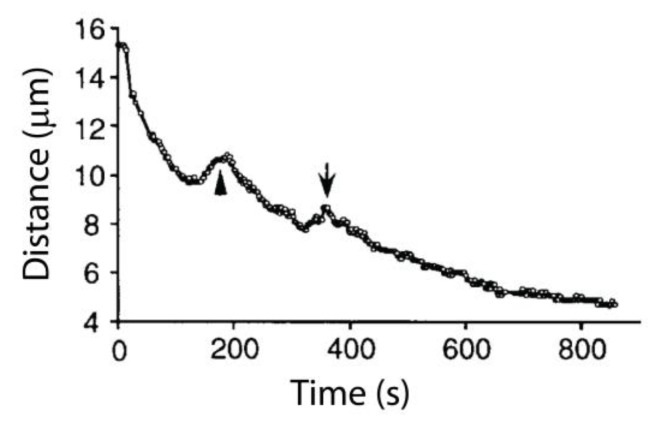
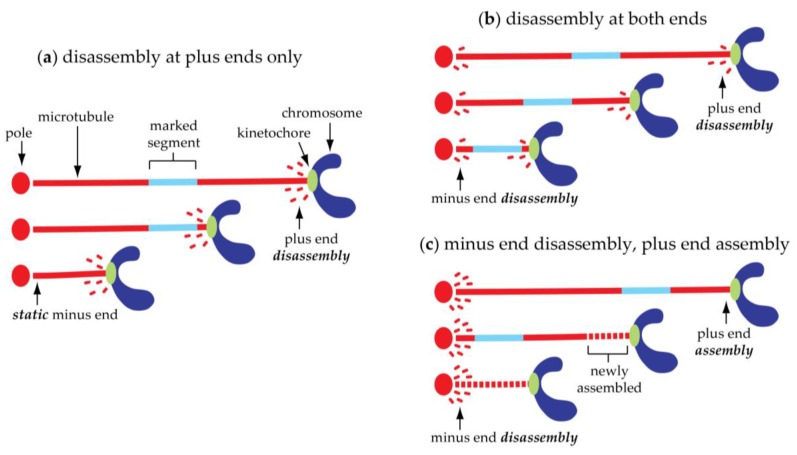
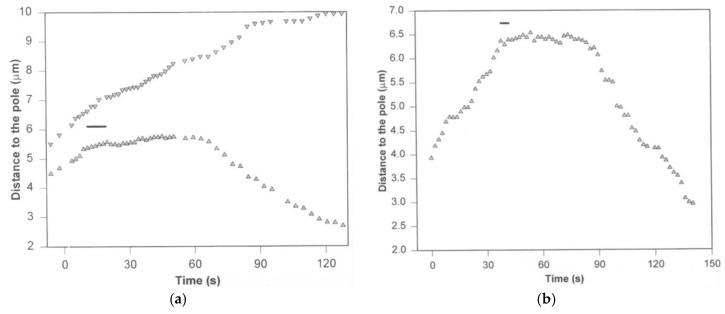
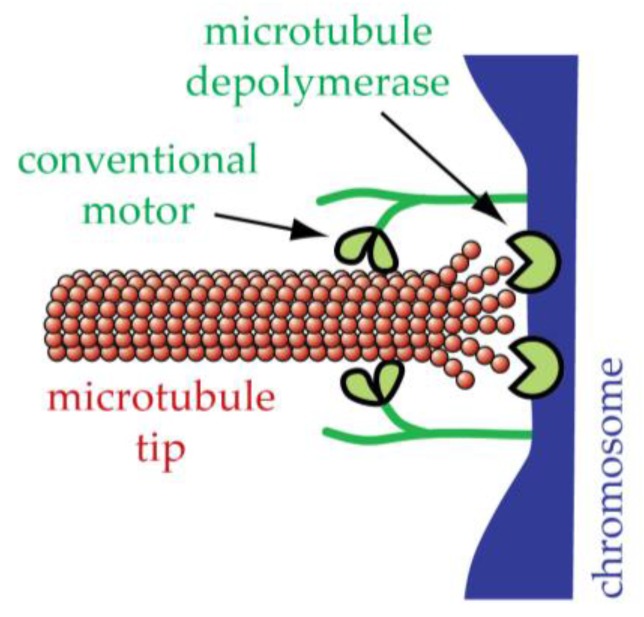
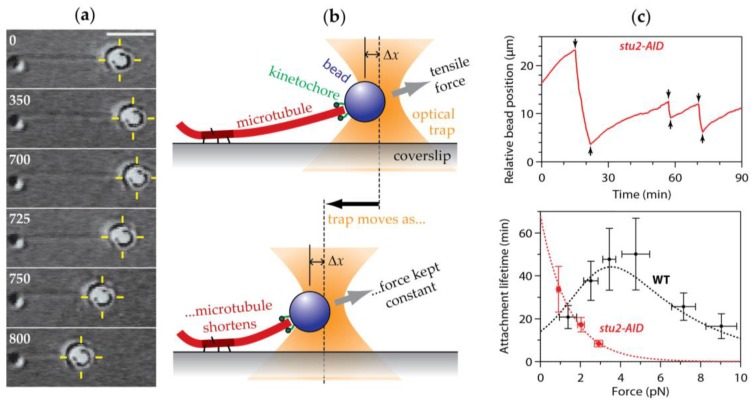
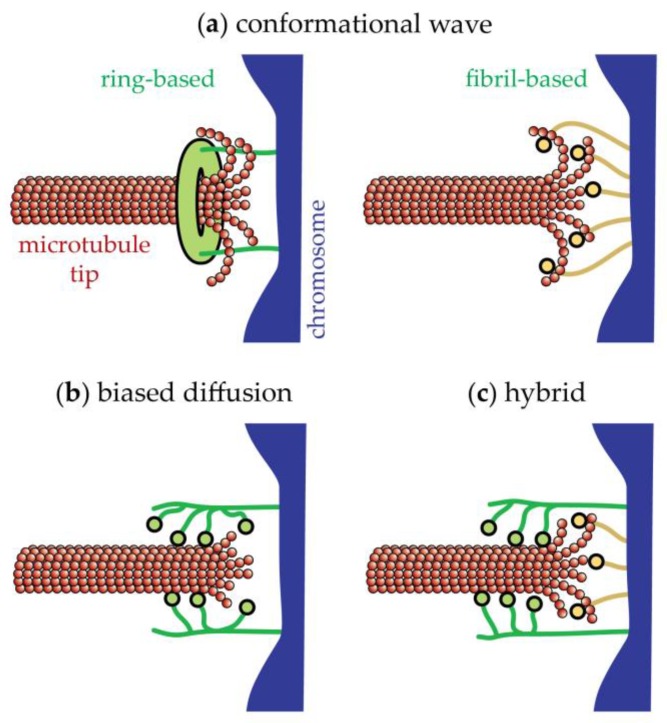
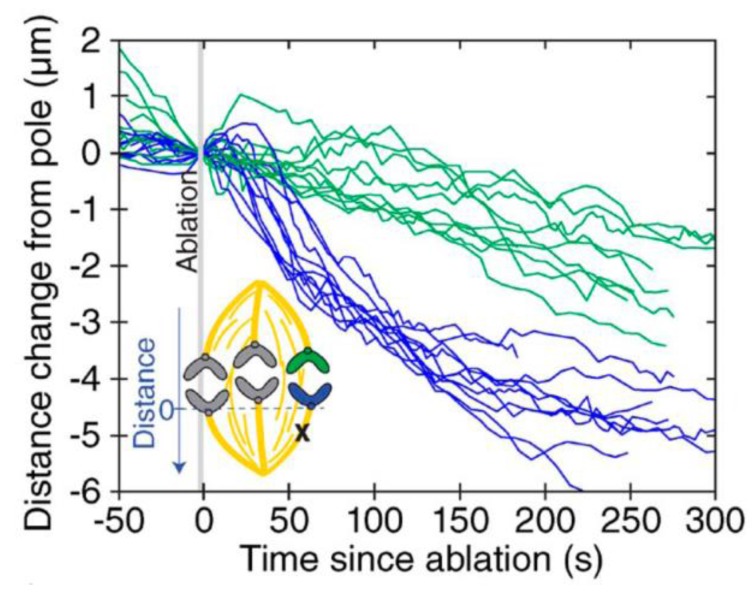
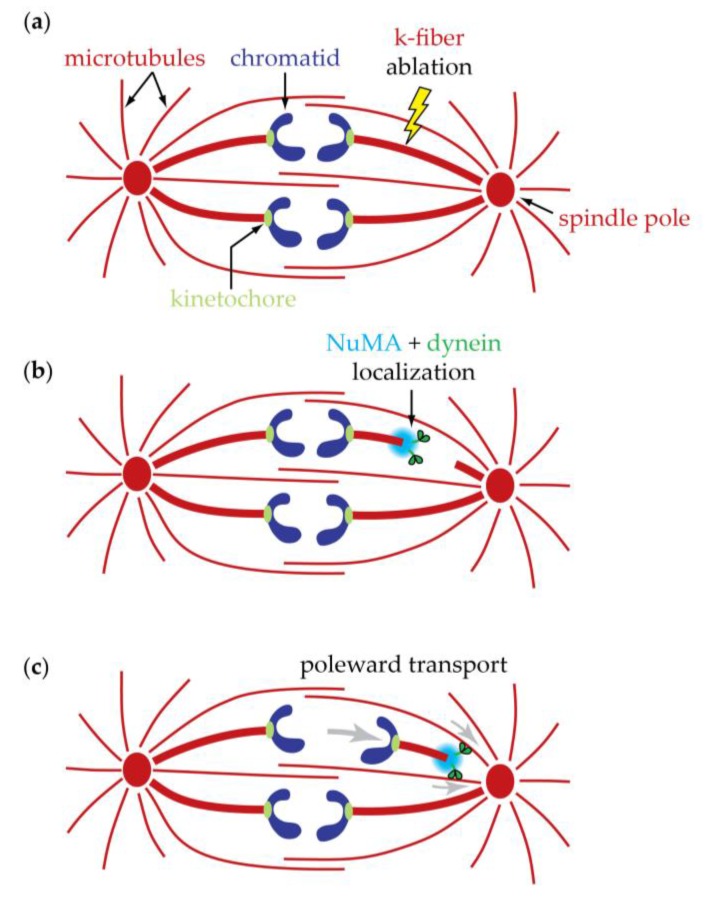
The separation of sister chromatids during anaphase is the culmination of mitosis and one of the most strikingly beautiful examples of cellular movement. It consists of two distinct processes: Anaphase A, the movement of chromosomes toward spindle poles via shortening of the connecting fibers, and anaphase B, separation of the two poles from one another via spindle elongation. I focus here on anaphase A chromosome-to-pole movement. The chapter begins by summarizing classical observations of chromosome movements, which support the current understanding of anaphase mechanisms. Live cell fluorescence microscopy studies showed that poleward chromosome movement is associated with disassembly of the kinetochore-attached microtubule fibers that link chromosomes to poles. Microtubule-marking techniques established that kinetochore-fiber disassembly often occurs through loss of tubulin subunits from the kinetochore-attached plus ends. In addition, kinetochore-fiber disassembly in many cells occurs partly through 'flux', where the microtubules flow continuously toward the poles and tubulin subunits are lost from minus ends. Molecular mechanistic models for how load-bearing attachments are maintained to disassembling microtubule ends, and how the forces are generated to drive these disassembly-coupled movements, are discussed.
Keywords: anaphase A; biased diffusion; chromosome-to-pole motion; conformational wave; kinetochore; microtubule poleward flux.
Conflict of interest statement
The author declares no conflict of interest.
Figures
References
-
- Mazia D. Mitosis and the physiology of cell division. In: Brachet J., Mirsky A.E., editors. The Cell: Biochemistry, Physiology, Morphology, Vol III. Academic Press; London, UK: 1961. pp. 77–412.
-
- Strasburger E. Die Controversen der indirecten Kerntheilung. Arch. Mikrosk. Anat. 1884;23:246–304. doi: 10.1007/BF02952312. - DOI
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
