

MHC-I peptides get out of the groove and enable a novel mechanism of HIV-1 escape
- PMID: 28218747
- PMCID: PMC7900914
- DOI: 10.1038/nsmb.3381
MHC-I peptides get out of the groove and enable a novel mechanism of HIV-1 escape
Abstract
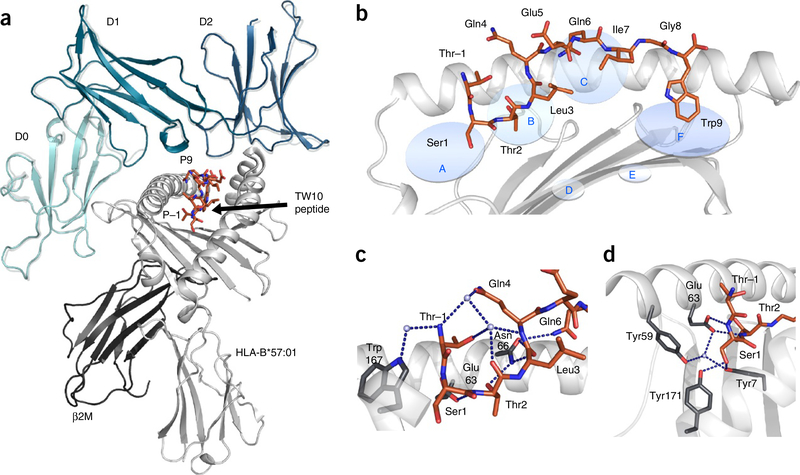
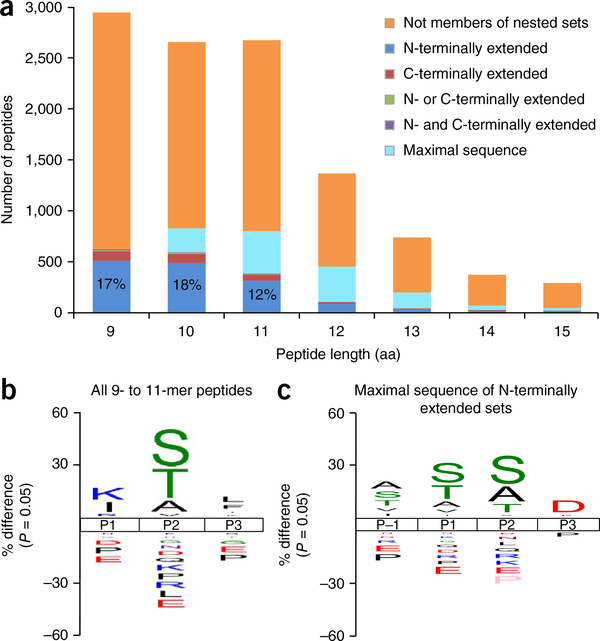
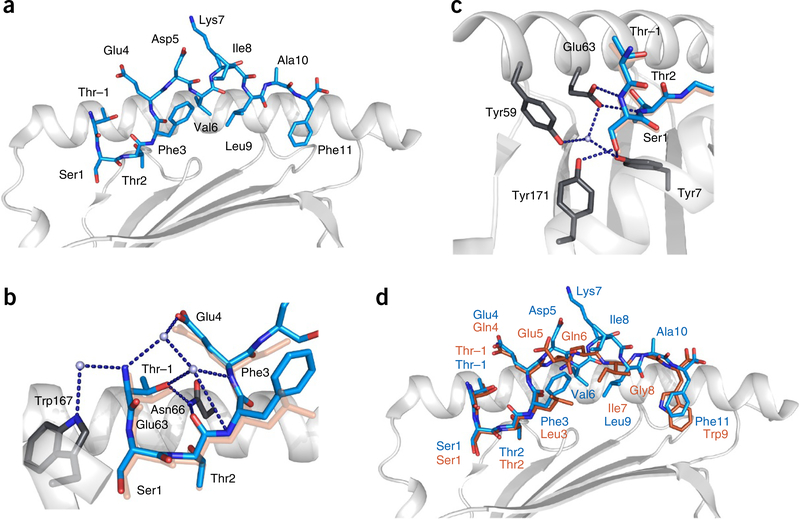
Major histocompatibility complex class I (MHC-I) molecules play a crucial role in immunity by capturing peptides for presentation to T cells and natural killer (NK) cells. The peptide termini are tethered within the MHC-I antigen-binding groove, but it is unknown whether other presentation modes occur. Here we show that 20% of the HLA-B*57:01 peptide repertoire comprises N-terminally extended sets characterized by a common motif at position 1 (P1) to P2. Structures of HLA-B*57:01 presenting N-terminally extended peptides, including the immunodominant HIV-1 Gag epitope TW10 (TSTLQEQIGW), showed that the N terminus protrudes from the peptide-binding groove. The common escape mutant TSNLQEQIGW bound HLA-B*57:01 canonically, adopting a dramatically different conformation than the TW10 peptide. This affected recognition by killer cell immunoglobulin-like receptor (KIR) 3DL1 expressed on NK cells. We thus define a previously uncharacterized feature of the human leukocyte antigen class I (HLA-I) immunopeptidome that has implications for viral immune escape. We further suggest that recognition of the HLA-B*57:01-TW10 epitope is governed by a 'molecular tension' between the adaptive and innate immune systems.
Conflict of interest statement
COMPETING FINANCIAL INTERESTS
The authors declare no competing financial interests.
Figures



References
-
- Saper MA, Bjorkman PJ & Wiley DC Refined structure of the human histocompatibility antigen HLA-A2 at 2.6 Å resolution. J. Mol. Biol. 219, 277–319 (1991). - PubMed
-
- Rammensee HG, Friede T & Stevanoviíc S MHC ligands and peptide motifs: first listing. Immunogenetics 41, 178–228 (1995). - PubMed
-
- Deres K, Beck W, Faath S, Jung G & Rammensee HG MHC/peptide binding studies indicate hierarchy of anchor residues. Cell. Immunol. 151, 158–167 (1993). - PubMed
-
- Wilson IA & Fremont DH Structural analysis of MHC class I molecules with bound peptide antigens. Semin. Immunol. 5, 75–80 (1993). - PubMed
-
- Garrett TP, Saper MA, Bjorkman PJ, Strominger JL & Wiley DC Specificity pockets for the side chains of peptide antigens in HLA-Aw68. Nature 342, 692–696 (1989). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
