

Dynamic protein S-palmitoylation mediates parasite life cycle progression and diverse mechanisms of virulence
- PMID: 28228066
- PMCID: PMC5560270
- DOI: 10.1080/10409238.2017.1287161
Dynamic protein S-palmitoylation mediates parasite life cycle progression and diverse mechanisms of virulence
Abstract
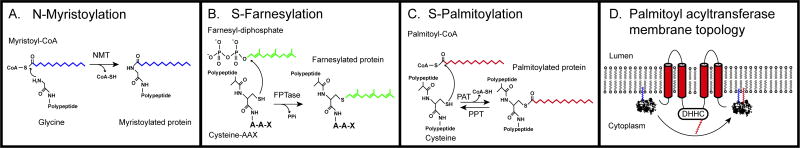
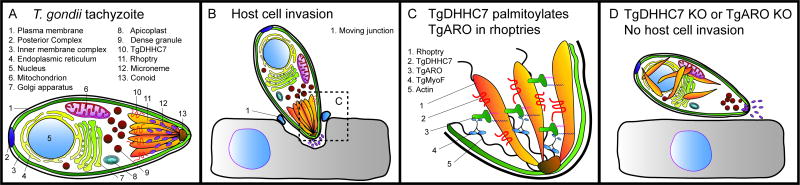
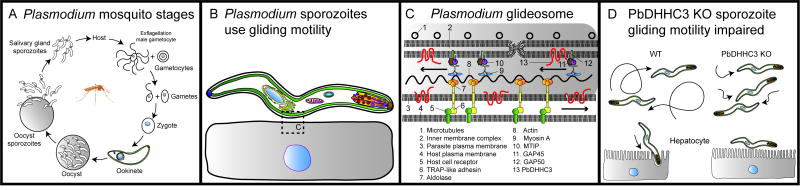
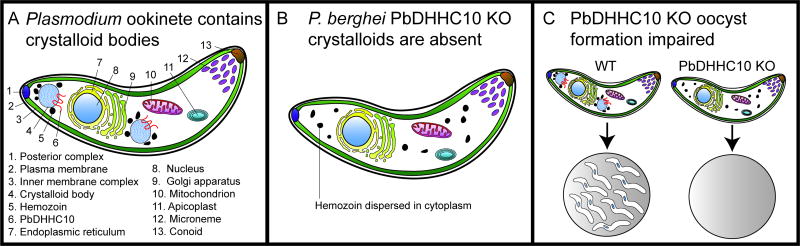
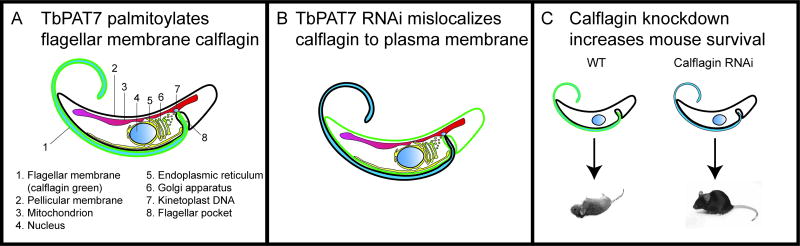
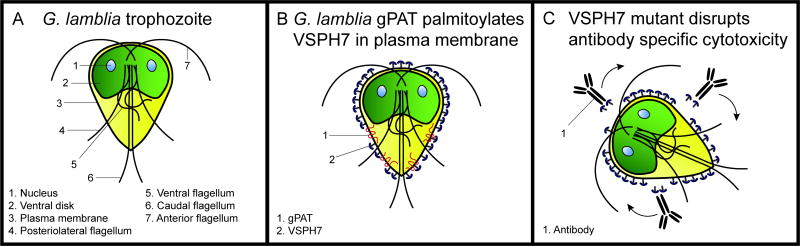
Eukaryotic parasites possess complex life cycles and utilize an assortment of molecular mechanisms to overcome physical barriers, suppress and/or bypass the host immune response, including invading host cells where they can replicate in a protected intracellular niche. Protein S-palmitoylation is a dynamic post-translational modification in which the fatty acid palmitate is covalently linked to cysteine residues on proteins by the enzyme palmitoyl acyltransferase (PAT) and can be removed by lysosomal palmitoyl-protein thioesterase (PPT) or cytosolic acyl-protein thioesterase (APT). In addition to anchoring proteins to intracellular membranes, functions of dynamic palmitoylation include - targeting proteins to specific intracellular compartments via trafficking pathways, regulating the cycling of proteins between membranes, modulating protein function and regulating protein stability. Recent studies in the eukaryotic parasites - Plasmodium falciparum, Toxoplasma gondii, Trypanosoma brucei, Cryptococcus neoformans and Giardia lamblia - have identified large families of PATs and palmitoylated proteins. Many palmitoylated proteins are important for diverse aspects of pathogenesis, including differentiation into infective life cycle stages, biogenesis and tethering of secretory organelles, assembling the machinery powering motility and targeting virulence factors to the plasma membrane. This review aims to summarize our current knowledge of palmitoylation in eukaryotic parasites, highlighting five exemplary mechanisms of parasite virulence dependent on palmitoylation.
Keywords: Giardia; Palmitoylation; Plasmodium; Toxoplasma; Trypanosoma; palmitoyl acyltransferase; pathogenesis.
Conflict of interest statement
The authors report no conflicts of interest.
Figures
References
-
- Aicart-Ramos C, Valero RA, Rodriguez-Crespo I. Protein palmitoylation and subcellular trafficking. Biochim Biophys Acta. 2011;1808:2981–94. - PubMed
-
- Albisetti A, Wiese S, Schneider A, Niemann M. A component of the mitochondrial outer membrane proteome of T. brucei probably contains covalent bound fatty acids. Exp Parasitol. 2015;155:49–57. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources