

E3 Ligase RNF126 Directly Ubiquitinates Frataxin, Promoting Its Degradation: Identification of a Potential Therapeutic Target for Friedreich Ataxia
- PMID: 28228265
- PMCID: PMC5329121
- DOI: 10.1016/j.celrep.2017.01.079
E3 Ligase RNF126 Directly Ubiquitinates Frataxin, Promoting Its Degradation: Identification of a Potential Therapeutic Target for Friedreich Ataxia
Abstract
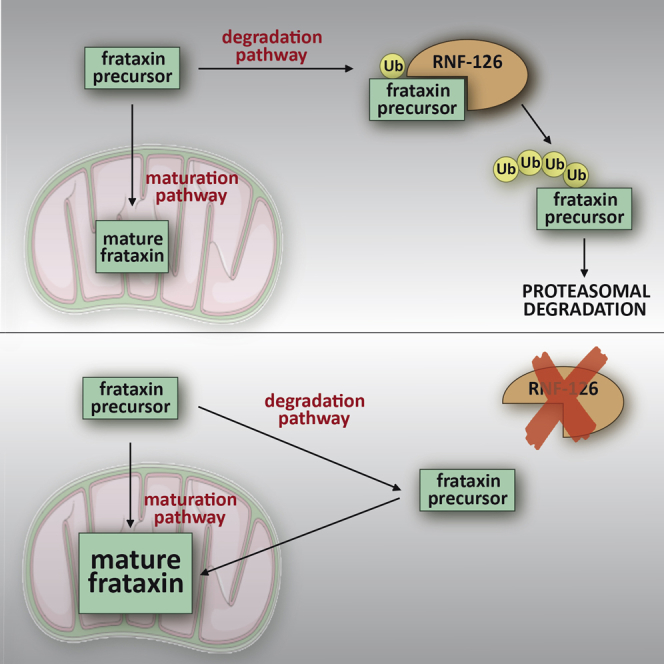
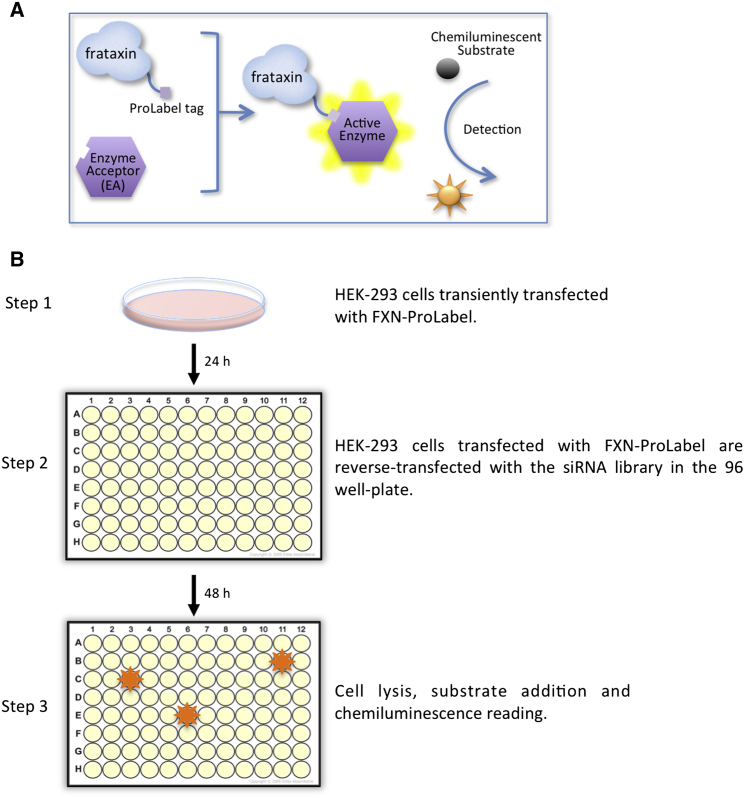
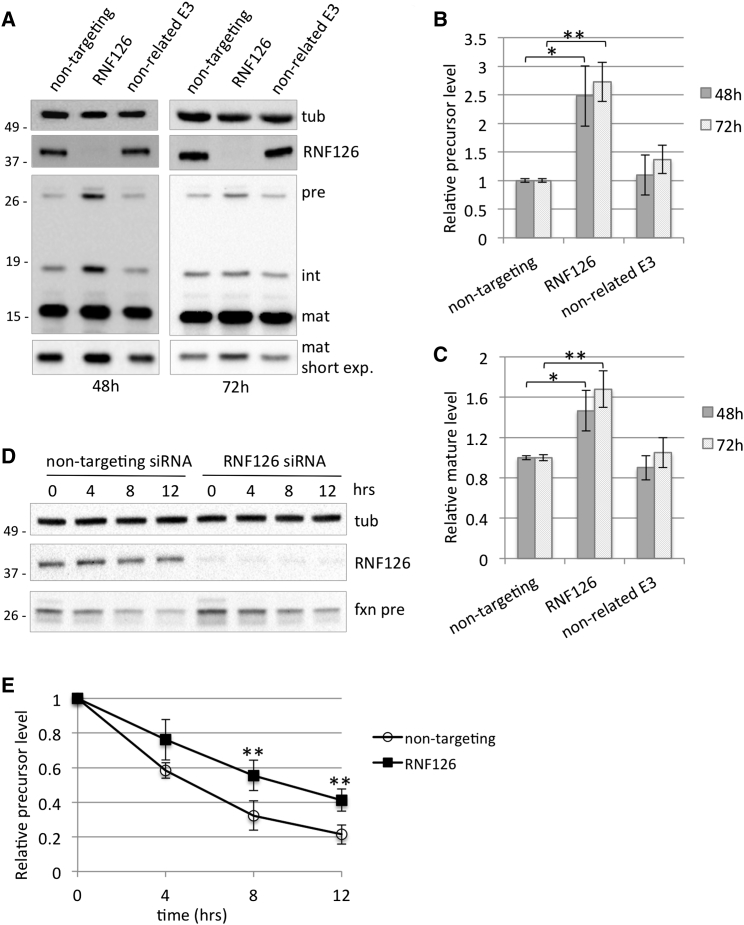
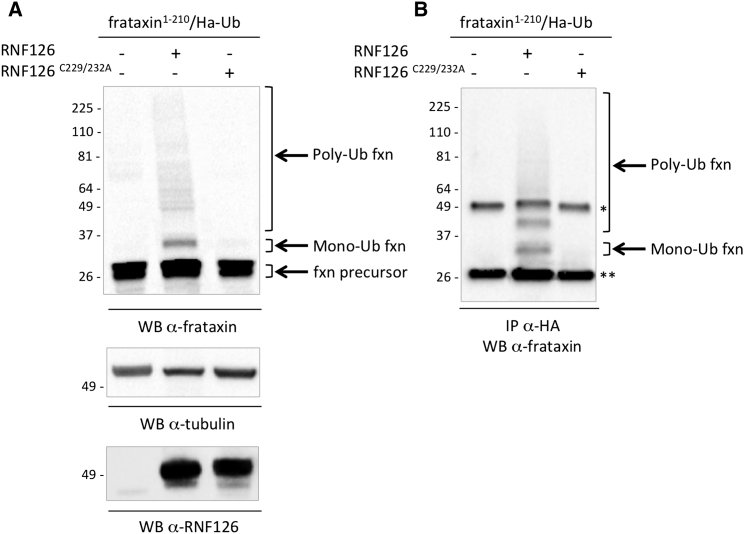
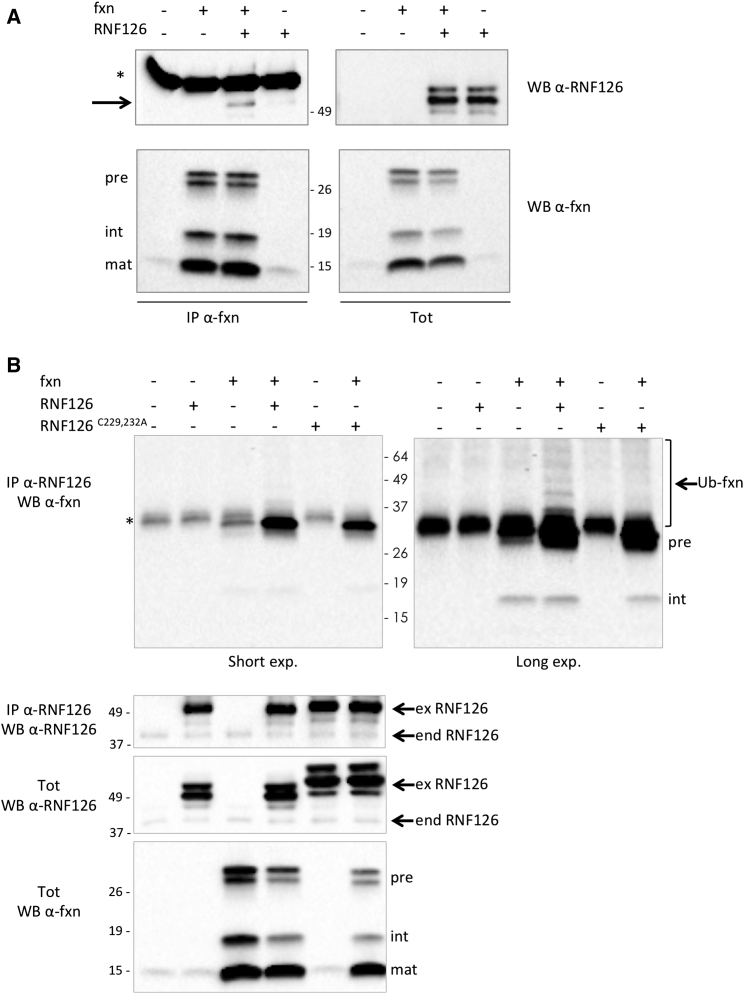
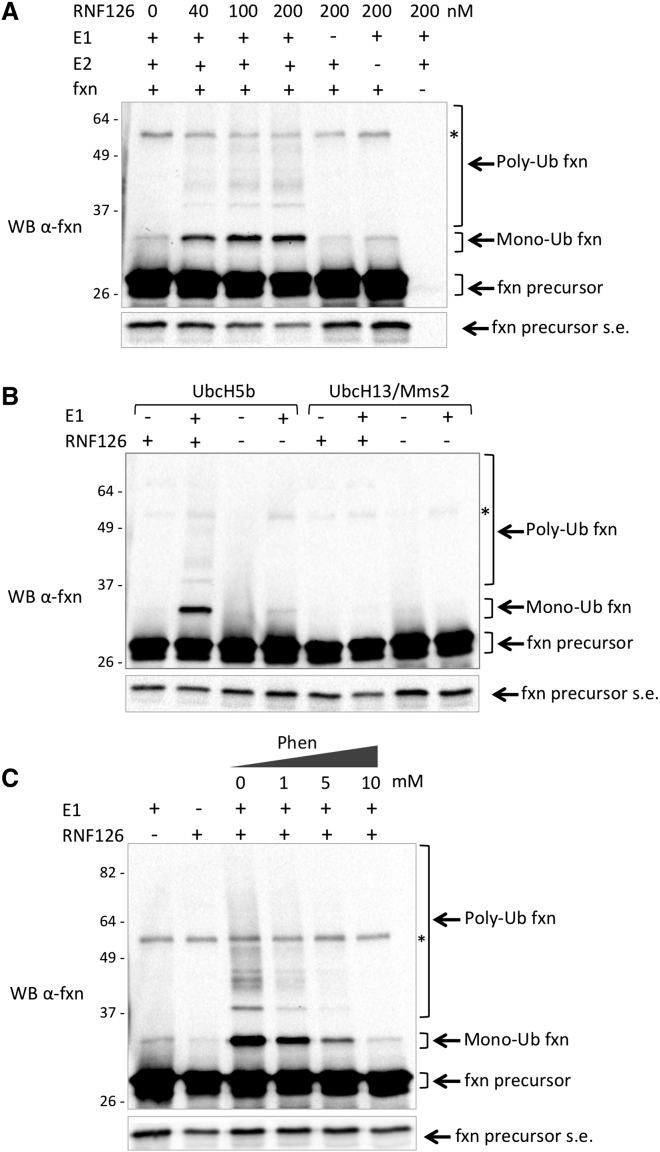
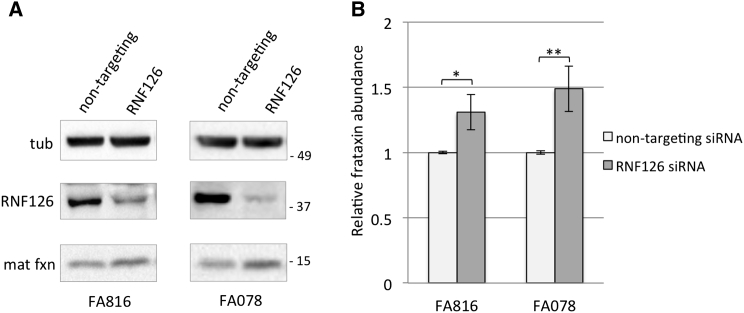
Friedreich ataxia (FRDA) is a severe genetic neurodegenerative disease caused by reduced expression of the mitochondrial protein frataxin. To date, there is no therapy to treat this condition. The amount of residual frataxin critically affects the severity of the disease; thus, attempts to restore physiological frataxin levels are considered therapeutically relevant. Frataxin levels are controlled by the ubiquitin-proteasome system; therefore, inhibition of the frataxin E3 ligase may represent a strategy to achieve an increase in frataxin levels. Here, we report the identification of the RING E3 ligase RNF126 as the enzyme that specifically mediates frataxin ubiquitination and targets it for degradation. RNF126 interacts with frataxin and promotes its ubiquitination in a catalytic activity-dependent manner, both in vivo and in vitro. Most importantly, RNF126 depletion results in frataxin accumulation in cells derived from FRDA patients, highlighting the relevance of RNF126 as a new therapeutic target for Friedreich ataxia.
Keywords: E3 ligase; Friedreich ataxia; RNF126; frataxin; protein degradation; therapeutic target; ubiquitin.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Preventing the ubiquitin-proteasome-dependent degradation of frataxin, the protein defective in Friedreich's ataxia.Hum Mol Genet. 2011 Apr 1;20(7):1253-61. doi: 10.1093/hmg/ddq566. Epub 2011 Jan 7. Hum Mol Genet. 2011. PMID: 21216878
-
Highly specific ubiquitin-competing molecules effectively promote frataxin accumulation and partially rescue the aconitase defect in Friedreich ataxia cells.Neurobiol Dis. 2015 Mar;75:91-9. doi: 10.1016/j.nbd.2014.12.011. Epub 2014 Dec 27. Neurobiol Dis. 2015. PMID: 25549872 Free PMC article.
-
GRP75 overexpression rescues frataxin deficiency and mitochondrial phenotypes in Friedreich ataxia cellular models.Hum Mol Genet. 2019 May 15;28(10):1594-1607. doi: 10.1093/hmg/ddy448. Hum Mol Genet. 2019. PMID: 30590615 Free PMC article.
-
Targeting the gene in Friedreich ataxia.Biochimie. 2008 Aug;90(8):1131-9. doi: 10.1016/j.biochi.2007.12.005. Epub 2007 Dec 28. Biochimie. 2008. PMID: 18206656 Review.
-
Friedreich ataxia: an update on animal models, frataxin function and therapies.Adv Exp Med Biol. 2009;652:247-61. doi: 10.1007/978-90-481-2813-6_17. Adv Exp Med Biol. 2009. PMID: 20225031 Review.
Cited by
-
The Ubiquitin Proteasome System in Neuromuscular Disorders: Moving Beyond Movement.Int J Mol Sci. 2020 Sep 3;21(17):6429. doi: 10.3390/ijms21176429. Int J Mol Sci. 2020. PMID: 32899400 Free PMC article. Review.
-
E3 ubiquitin ligases and deubiquitinases in bladder cancer tumorigenesis and implications for immunotherapies.Front Immunol. 2023 Jul 11;14:1226057. doi: 10.3389/fimmu.2023.1226057. eCollection 2023. Front Immunol. 2023. PMID: 37497216 Free PMC article. Review.
-
Ubiquitylomics: An Emerging Approach for Profiling Protein Ubiquitylation in Skeletal Muscle.J Cachexia Sarcopenia Muscle. 2024 Dec;15(6):2281-2294. doi: 10.1002/jcsm.13601. Epub 2024 Sep 16. J Cachexia Sarcopenia Muscle. 2024. PMID: 39279720 Free PMC article. Review.
-
High Expression of RING Finger Protein 126 Predicts Unfavorable Prognosis of Epithelial Ovarian Cancer.Med Sci Monit. 2020 Apr 7;26:e921370. doi: 10.12659/MSM.921370. Med Sci Monit. 2020. PMID: 32254065 Free PMC article.
-
Molecular Mechanisms and Therapeutics for the GAA·TTC Expansion Disease Friedreich Ataxia.Neurotherapeutics. 2019 Oct;16(4):1032-1049. doi: 10.1007/s13311-019-00764-x. Neurotherapeutics. 2019. PMID: 31317428 Free PMC article. Review.
References
-
- Al-Mahdawi S., Pinto R.M., Ismail O., Varshney D., Lymperi S., Sandi C., Trabzuni D., Pook M. The Friedreich ataxia GAA repeat expansion mutation induces comparable epigenetic changes in human and transgenic mouse brain and heart tissues. Hum. Mol. Genet. 2008;17:735–746. - PubMed
-
- Alper G., Narayanan V. Friedreich’s ataxia. Pediatr. Neurol. 2003;28:335–341. - PubMed
-
- Berndsen C.E., Wolberger C. New insights into ubiquitin E3 ligase mechanism. Nat. Struct. Mol. Biol. 2014;21:301–307. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
