

Proofreading of Peptide-MHC Complexes through Dynamic Multivalent Interactions
- PMID: 28228754
- PMCID: PMC5296336
- DOI: 10.3389/fimmu.2017.00065
Proofreading of Peptide-MHC Complexes through Dynamic Multivalent Interactions
Abstract
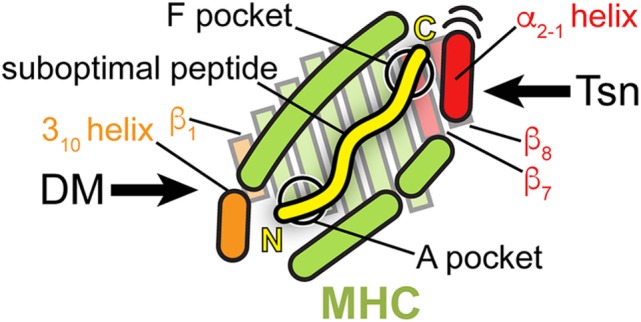
The adaptive immune system is able to detect and destroy cells that are malignantly transformed or infected by intracellular pathogens. Specific immune responses against these cells are elicited by antigenic peptides that are presented on major histocompatibility complex class I (MHC I) molecules and recognized by cytotoxic T lymphocytes at the cell surface. Since these MHC I-presented peptides are generated in the cytosol by proteasomal protein degradation, they can be metaphorically described as a window providing immune cells with insights into the state of the cellular proteome. A crucial element of MHC I antigen presentation is the peptide-loading complex (PLC), a multisubunit machinery, which contains as key constituents the transporter associated with antigen processing (TAP) and the MHC I-specific chaperone tapasin (Tsn). While TAP recognizes and shuttles the cytosolic antigenic peptides into the endoplasmic reticulum (ER), Tsn samples peptides in the ER for their ability to form stable complexes with MHC I, a process called peptide proofreading or peptide editing. Through its selection of peptides that improve MHC I stability, Tsn contributes to the hierarchy of immunodominant peptide epitopes. Despite the fact that it concerns a key event in adaptive immunity, insights into the catalytic mechanism of peptide proofreading carried out by Tsn have only lately been gained via biochemical, biophysical, and structural studies. Furthermore, a Tsn homolog called TAP-binding protein-related (TAPBPR) has only recently been demonstrated to function as a second MHC I-specific chaperone and peptide proofreader. Although TAPBPR is PLC-independent and has a distinct allomorph specificity, it is likely to share a common catalytic mechanism with Tsn. This review focuses on the current knowledge of the multivalent protein-protein interactions and the concomitant dynamic molecular processes underlying peptide-proofreading catalysis. We do not only derive a model that highlights the common mechanistic principles shared by the MHC I editors Tsn and TAPBPR, and the MHC II editor HLA-DM, but also illustrate the distinct quality control strategies employed by these chaperones to sample epitopes. Unraveling the mechanistic underpinnings of catalyzed peptide proofreading will be crucial for a thorough understanding of many aspects of immune recognition, from infection control and tumor immunity to autoimmune diseases and transplant rejection.
Keywords: MHC; adaptive immunity; antigen presentation; molecular tug-of-war; peptide editing; peptide-loading complex; quality control; tapasin.
Figures




References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous