

Trichoderma reesei xylanase 5 is defective in the reference strain QM6a but functional alleles are present in other wild-type strains
- PMID: 28229208
- PMCID: PMC5403845
- DOI: 10.1007/s00253-017-8161-4
Trichoderma reesei xylanase 5 is defective in the reference strain QM6a but functional alleles are present in other wild-type strains
Abstract
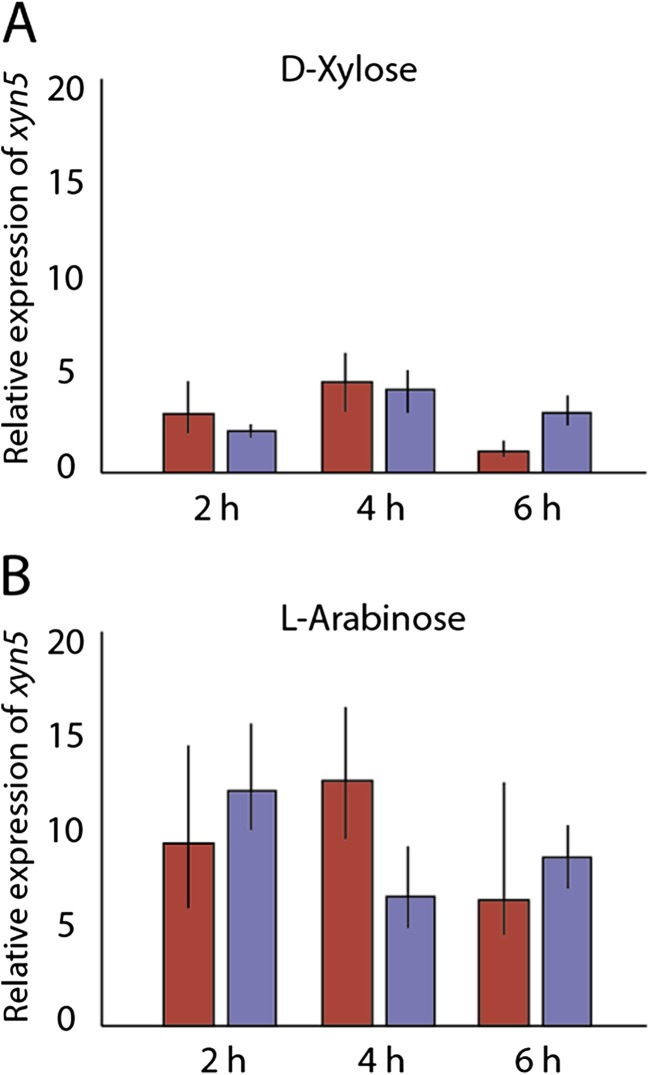
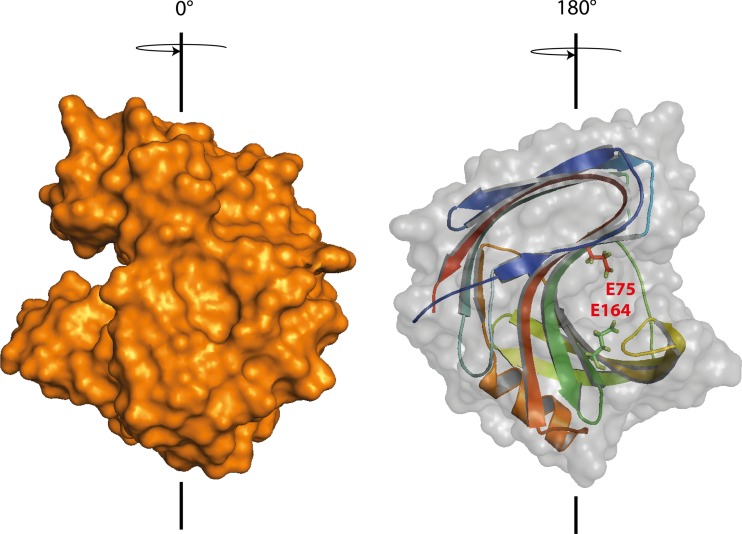
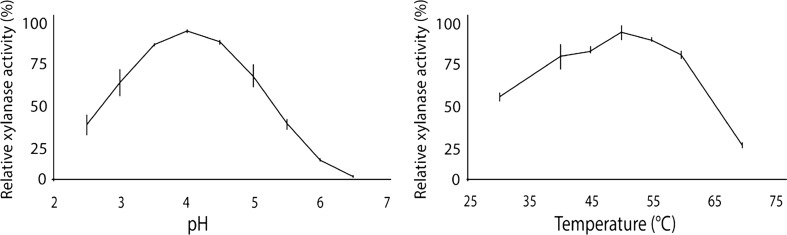
Trichoderma reesei is a paradigm for the regulation and industrial production of plant cell wall-degrading enzymes. Among these, five xylanases, including the glycoside hydrolase (GH) family 11 XYN1 and XYN2, the GH10 XYN3, and the GH30 XYN4 and XYN6, were described. By genome mining and transcriptome analysis, a further putative xylanase, encoded by xyn5, was identified. Analysis of xyn5 from the genome-sequenced reference strain T. reesei QM6a shows that it encodes a non-functional, truncated form of XYN5. However, non-truncated orthologues are present in other genome sequenced Trichoderma spp., and sequencing of xyn5 in other T. reesei wild-type isolates shows that they harbor a putative functional xyn5 allele. In silico analysis and 3D modeling revealed that the encoded XYN5 has significant structural similarities to xylanases of the GH11 family, including a GH-typical substrate binding groove and a carboxylate pair in the active site. The xyn5 of wild-type strain TUCIM1282 was recombinantly expressed in a T. reesei strain with a (hemi)cellulase-free background and the corresponding protein purified to apparent homogeneity. The pH and temperature optima and the kinetic parameters of the purified XYN5 were pH 4, 50 °C, and V max = 2646 nkat/mg with a K m of 9.68 mg/ml. This functional xyn5 allele was used to replace the mutated version which led to an overall increase of the xylanolytic activity. These findings are of particular importance as GH11 xylanases are of high biotechnological relevance, and T. reesei is one of the main industrial producers of such lignocellulose-degrading enzymes.
Keywords: Biofuels; Biorefinery; GH11 family; Recombinant protein production; Trichoderma reesei; XYN1; Xylanase.
Conflict of interest statement
Funding
This study was supported by grant P24219 of the Austrian Science Fund FWF to BS and JR. JR was co-financed by the TU Wien Ph.D. Program Molecular and Elemental Imaging in Biosciences MEIBio.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Figures



References
-
- Bailey M, Biely P, Poutanen K. Interlaboratory testing of methods for assay of xylanase activity. J Biotechnol. 1992;23:257–270. doi: 10.1016/0168-1656(92)90074-J. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
