

Simvastatin-induced cell cycle arrest through inhibition of STAT3/SKP2 axis and activation of AMPK to promote p27 and p21 accumulation in hepatocellular carcinoma cells
- PMID: 28230855
- PMCID: PMC5386458
- DOI: 10.1038/cddis.2016.472
Simvastatin-induced cell cycle arrest through inhibition of STAT3/SKP2 axis and activation of AMPK to promote p27 and p21 accumulation in hepatocellular carcinoma cells
Abstract
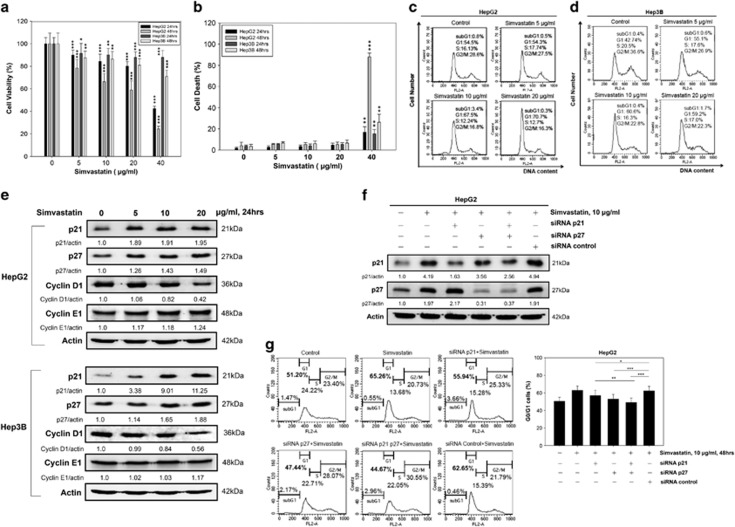
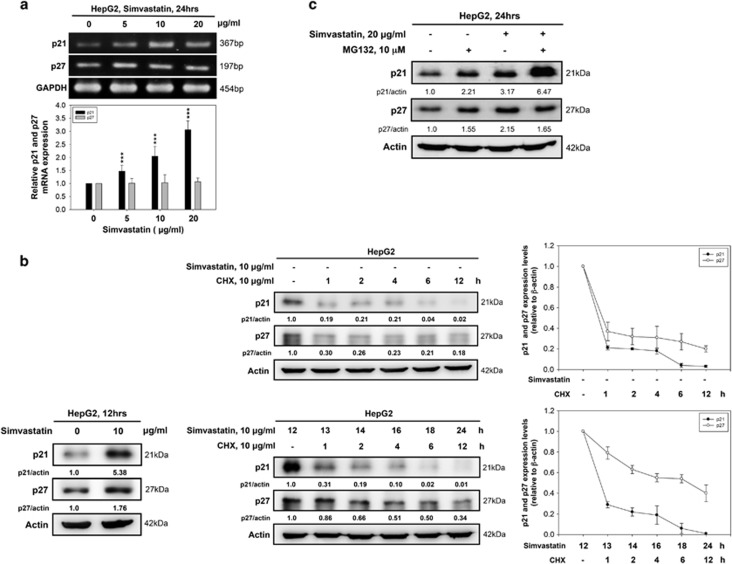
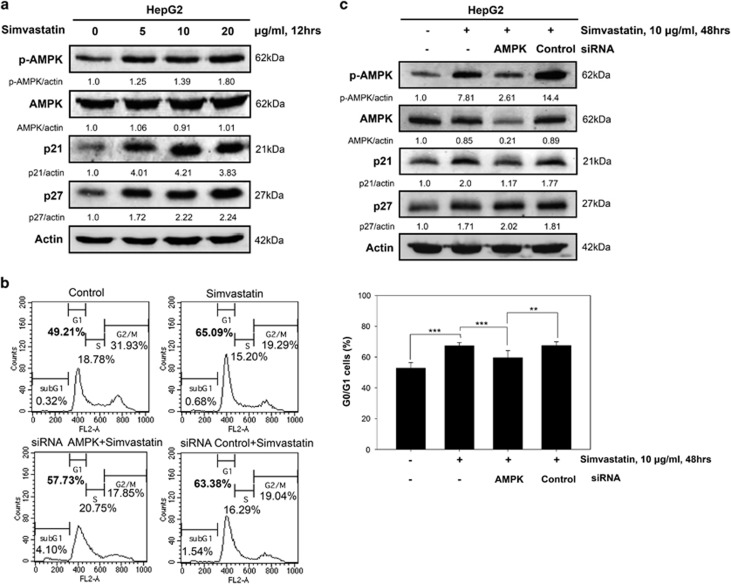
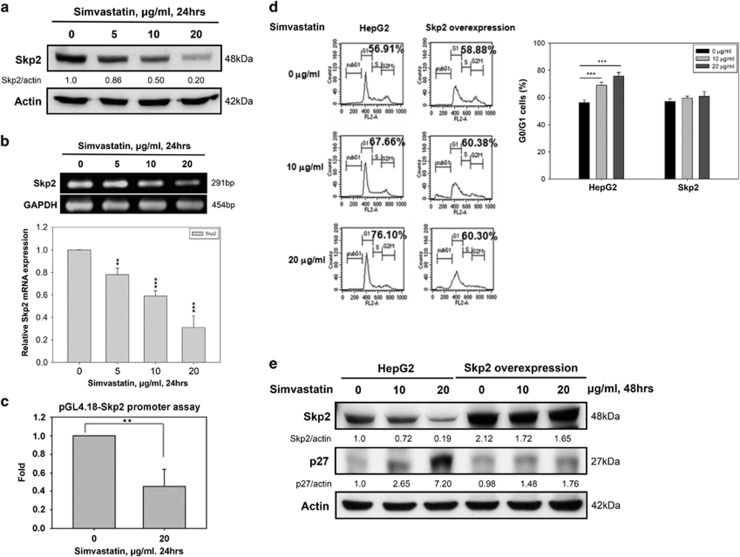
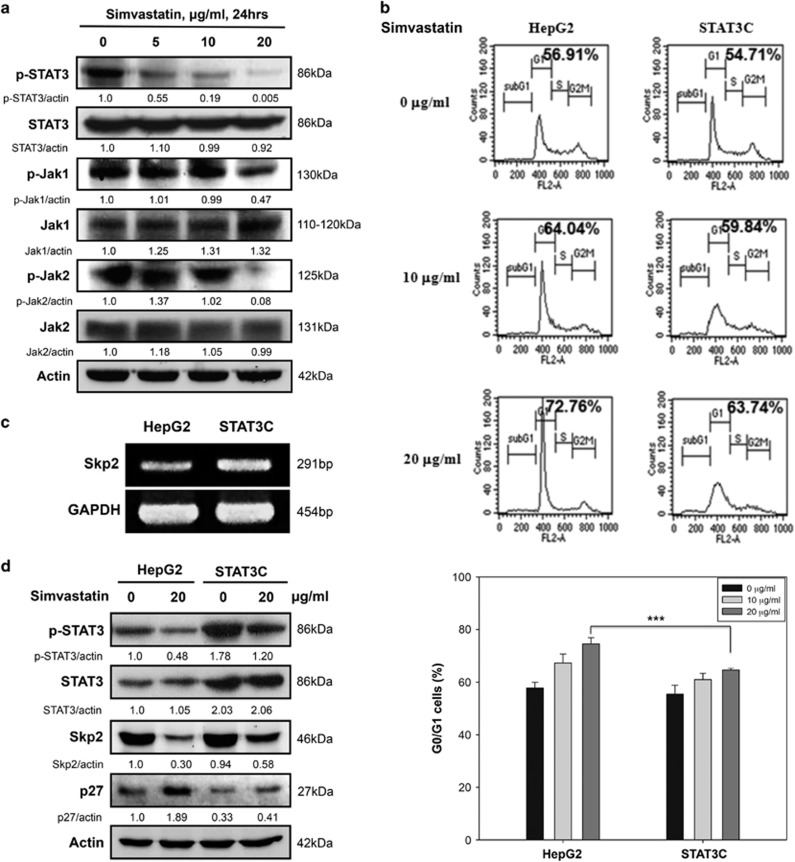
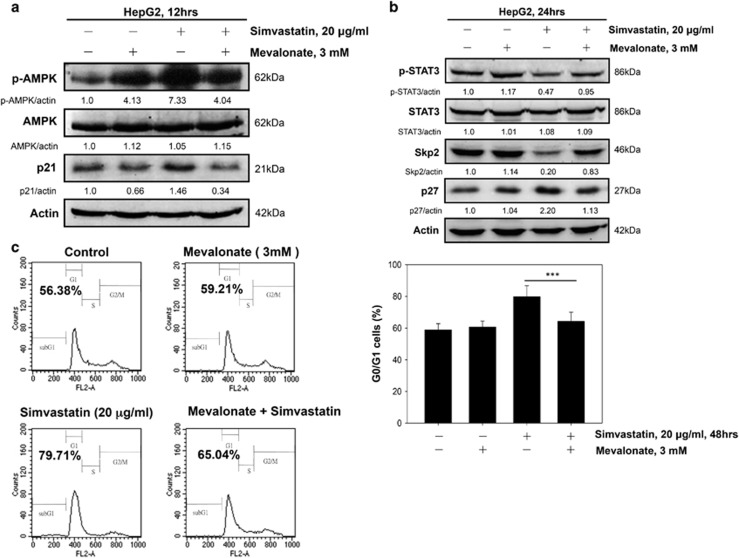
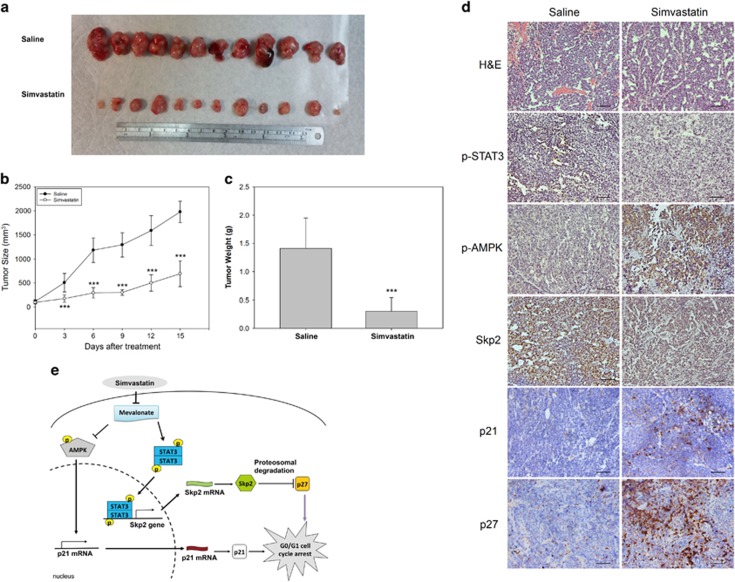
Hepatocellular carcinoma (HCC) is characterized by a poor prognosis and is one of the leading causes of cancer-related death worldwide. Simvastatin, an HMG-CoA reductase inhibitor, which decreases cholesterol synthesis by inhibiting mevalonate pathways and is widely used to treat cardiovascular diseases. Simvastatin exhibits anticancer effects against several malignancies. However, the molecular mechanisms underlying the anticancer effects of simvastatin on HCC are still not well understood. In this study, we demonstrated simvastatin-induced G0/G1 arrest by inducing p21 and p27 accumulation in HepG2 and Hep3B cells. Simvastatin also promoted AMP-activated protein kinase (AMPK) activation, which induced p21 upregulation by increasing its transcription. Consistent with this finding, we found genetic silencing of AMPK reduced p21 expression; however, AMPK silencing had no effect on p27 expression in HCC cells. Simvastatin decreased Skp2 expression at the transcriptional level, which resulted in p27 accumulation by preventing proteasomal degradation, an effect mediated by signal transducer and activator of transcription 3 (STAT3) inhibition. Constitutive STAT3 activation maintained high-level Skp2 expression and lower level p27 expression and significantly prevented G0/G1 arrest in simvastatin-treated HCC cells. Mevalonate decreased simvastatin-induced AMPK activation and rescued phospho-STAT3 and Skp2 expression in HCC cells, which resulted in the prevention of G0/G1 arrest through inhibition of p21 and p27 accumulation. Moreover, simvastatin significantly decreased tumor growth in HepG2 xenograft mice. Consistently, we found that simvastatin also increased p21 and p27 expression in tumor sections by reducing Skp2 expression and inducing AMPK activation and STAT3 suppression in the same tumor tissues. Taken together, these findings are demonstrative of the existence of a novel pathway in which simvastatin induces G0/G1 arrest by upregulating p21 and p27 by activating AMPK and inhibiting the STAT3-Skp2 axis, respectively. The results identify novel targets that explain the beneficial anticancer effects of simvastatin treatment on HCC in vitro and in vivo.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D et al. Global cancer statistics. CA Cancer J Clin 2011; 61: 69–90. - PubMed
-
- El-Serag HB, Rudolph KL. Hepatocellular carcinoma: epidemiology and molecular carcinogenesis. Gastroenterology 2007; 132: 2557–2576. - PubMed
-
- El-Serag HB. Hepatocellular carcinoma. N Engl J Med 2011; 365: 1118–1127. - PubMed
-
- Pang RW, Poon RT. From molecular biology to targeted therapies for hepatocellular carcinoma: the future is now. Oncology 2007; 72(Suppl 1): 30–44. - PubMed
-
- Wu CY, Chen YJ, Ho HJ, Hsu YC, Kuo KN, Wu MS et al. Association between nucleoside analogues and risk of hepatitis B virus-related hepatocellular carcinoma recurrence following liver resection. JAMA 2012; 308: 1906–1914. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
