

Precision engineering for PRRSV resistance in pigs: Macrophages from genome edited pigs lacking CD163 SRCR5 domain are fully resistant to both PRRSV genotypes while maintaining biological function
- PMID: 28231264
- PMCID: PMC5322883
- DOI: 10.1371/journal.ppat.1006206
Precision engineering for PRRSV resistance in pigs: Macrophages from genome edited pigs lacking CD163 SRCR5 domain are fully resistant to both PRRSV genotypes while maintaining biological function
Abstract
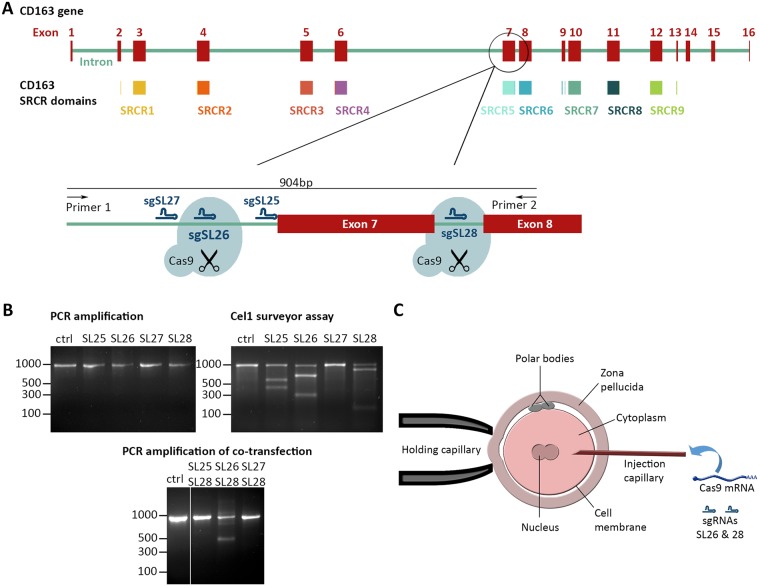
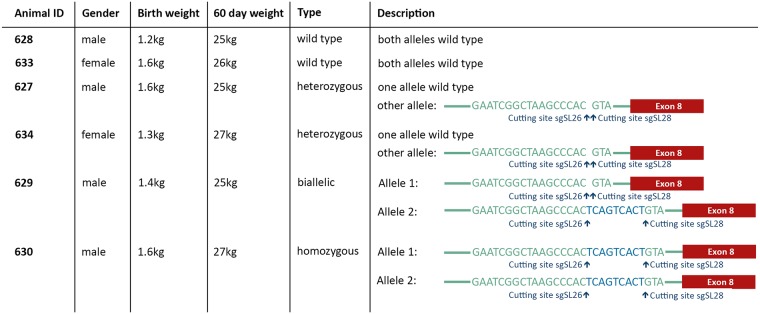
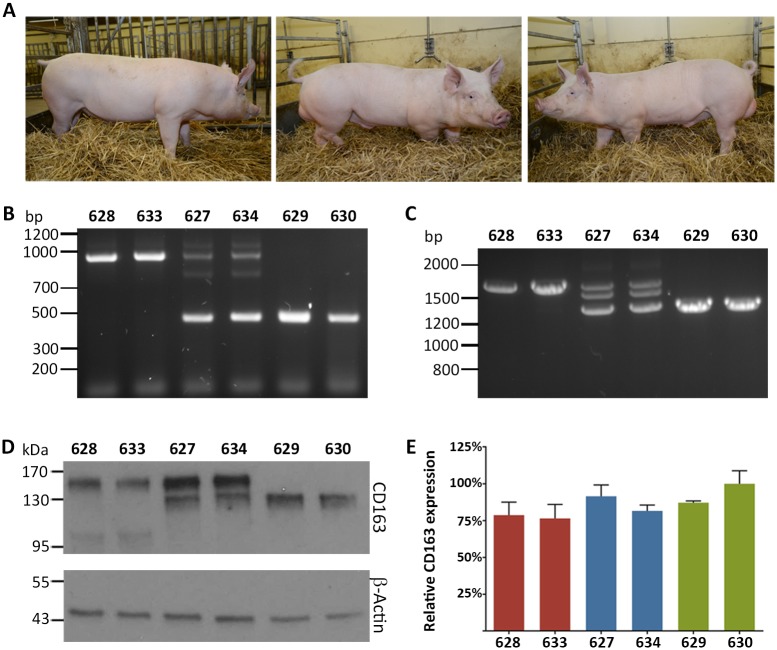
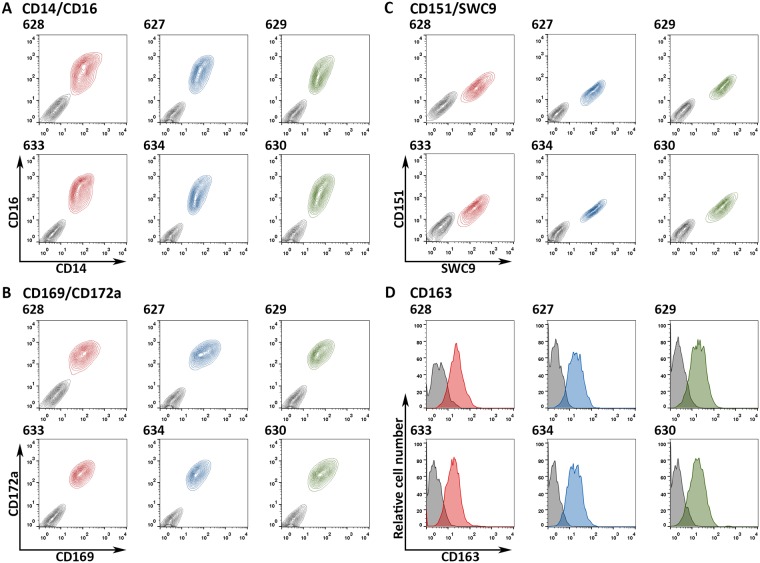
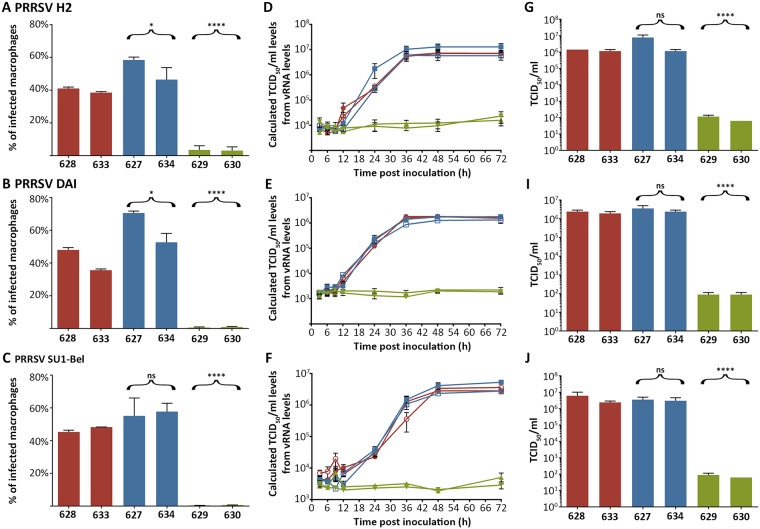
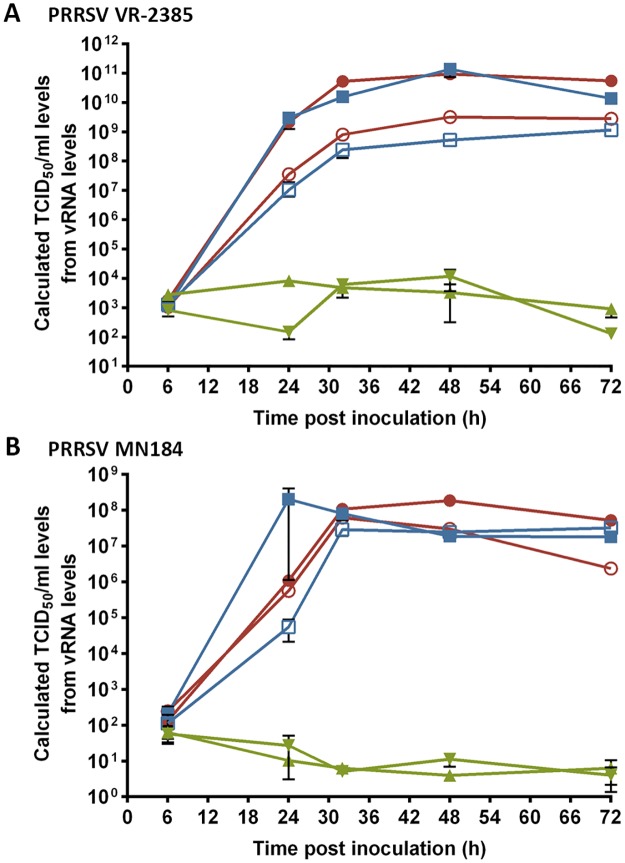
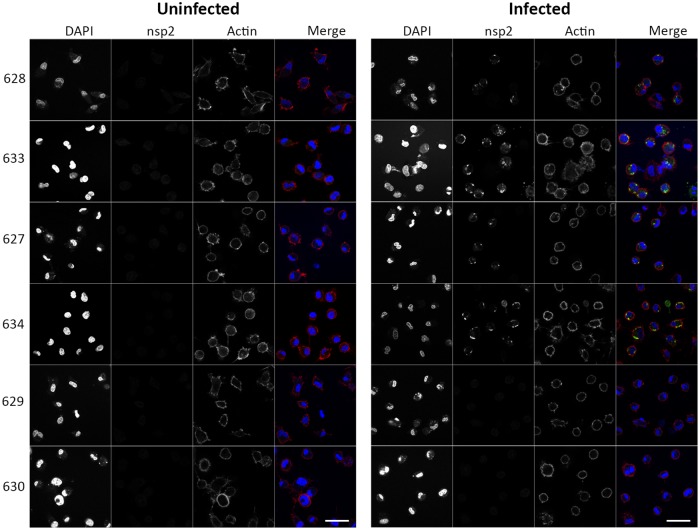
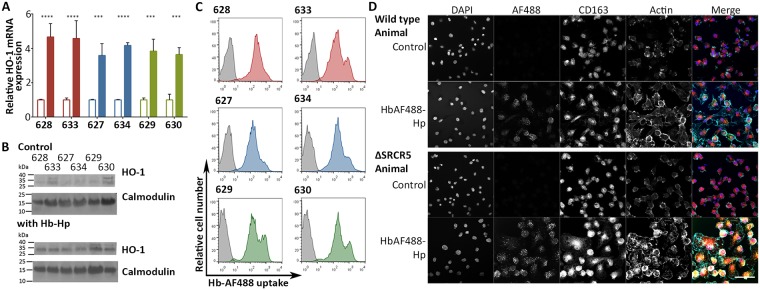
Porcine Reproductive and Respiratory Syndrome (PRRS) is a panzootic infectious disease of pigs, causing major economic losses to the world-wide pig industry. PRRS manifests differently in pigs of all ages but primarily causes late-term abortions and stillbirths in sows and respiratory disease in piglets. The causative agent of the disease is the positive-strand RNA PRRS virus (PRRSV). PRRSV has a narrow host cell tropism, limited to cells of the monocyte/macrophage lineage. CD163 has been described as a fusion receptor for PRRSV, whereby the scavenger receptor cysteine-rich domain 5 (SRCR5) region was shown to be an interaction site for the virus in vitro. CD163 is expressed at high levels on the surface of macrophages, particularly in the respiratory system. Here we describe the application of CRISPR/Cas9 to pig zygotes, resulting in the generation of pigs with a deletion of Exon 7 of the CD163 gene, encoding SRCR5. Deletion of SRCR5 showed no adverse effects in pigs maintained under standard husbandry conditions with normal growth rates and complete blood counts observed. Pulmonary alveolar macrophages (PAMs) and peripheral blood monocytes (PBMCs) were isolated from the animals and assessed in vitro. Both PAMs and macrophages obtained from PBMCs by CSF1 stimulation (PMMs) show the characteristic differentiation and cell surface marker expression of macrophages of the respective origin. Expression and correct folding of the SRCR5 deletion CD163 on the surface of macrophages and biological activity of the protein as hemoglobin-haptoglobin scavenger was confirmed. Challenge of both PAMs and PMMs with PRRSV genotype 1, subtypes 1, 2, and 3 and PMMs with PRRSV genotype 2 showed complete resistance to viral infections assessed by replication. Confocal microscopy revealed the absence of replication structures in the SRCR5 CD163 deletion macrophages, indicating an inhibition of infection prior to gene expression, i.e. at entry/fusion or unpacking stages.
Conflict of interest statement
I have read the journal's policy and the authors of this manuscript have the following competing interests: CB, SGL, TAA, CBAW, and ALA have patent claims on the targeted deletion described in the manuscript. AJM is employed by a commercial company, Genus plc, a commercial livestock breeder.
Figures
References
-
- Russell P, Atkinson K, Krishnan L (1980) Recurrent reproductive failure due to severe placental villitis of unknown etiology. J Reprod Med 24: 93–98. - PubMed
-
- Pejsak Z, Stadejek T, Markowska-Daniel I (1997) Clinical signs and economic losses caused by porcine reproductive and respiratory syndrome virus in a large breeding farm. Vet Microbiol 55: 317–322. - PubMed
-
- Christianson WT, Joo HS (1994) Porcine reproductive and respiratory syndrome: A review. J Swine Health Prod 2: 10–28.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
